
HLA-DQ1 is a serotype that covers a broad range of HLA-DQ haplotypes. Historically it was identified as a DR-like alpha chain called DC1; later, it was among 3 types DQw1, DQw2 and DQw3. Of these three serotyping specificities only DQw1 recognized DQ alpha chain. The serotype is positive in individuals who bear the DQA1*01 alleles. The most frequently found within this group are: DQA1*0101, *0102, *0103, and *0104. In the illustration on the right, DQ1 serotyping antibodies recognizes the DQ α (magenta), where antibodies to DQA1* gene products bind variable regions close to the peptide binding pocket.

HLA-DR2 (DR2) of the HLA-DR serotype system, is a broad antigen serotype that is now preferentially covered by HLA-DR15 and HLA-DR16 serotype group. This serotype primarily recognizes gene products of the HLA-DRB1*15 and HLA-DRB1*16 allele groups.

HLA-A*02 (A*02) is a human leukocyte antigen serotype within the HLA-A serotype group. The serotype is determined by the antibody recognition of the α2 domain of the HLA-A α-chain. For A*02, the α chain is encoded by the HLA-A*02 gene and the β chain is encoded by the B2M locus. In 2010 the World Health Organization Naming Committee for Factors of the HLA System revised the nomenclature for HLAs. Before this revision, HLA-A*02 was also referred to as HLA-A2, HLA-A02, and HLA-A*2.
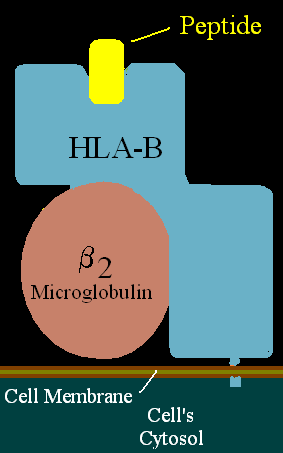
HLA-B*82 (B*82) is an HLA-B allele-group. There is no current useful serotyping for HLA-B*82 gene products. B*8201 was first identified by sequence analysis and appears to be derived by gene conversion between B*5602 and another HLA class I allele., later B*8202 was identified in a caucasian and was suggested to be ancestral to B*8201, as product between gene conversion of B*5602 allele and B*4501 allele. B*82 is more common in East Africa, Kenya and Sudan, the frequency of B*8201 is found in the peoples to the west, sporadically in Central and West Africa, B*8202 is found in Sudan and Saudi Arabia.
HLA-B78 (B78) is an HLA-B serotype. The serotype identifies the more common HLA-B*78 gene products. B78 is more common in West and North Africa, but is also scattered at low frequencies in parts of Asia.
HLA-B67 (B67) is an HLA-B serotype. The serotype identifies the more common HLA-B*67 gene products. B67 is region specific recombinant haplotype formed by the gene conversion of B*39, an allele common along the Northwest Pacific Rim, and B7, B22, or B27.
HLA-B59 (B59) is an HLA-B serotype. The serotype identifies the more common HLA-B*## gene products. B59 is a hybrid between B*55 and B*51. B59 is more common in Japan, Korea, N. China and Mongolia.
HLA-B47 (B47) is an HLA–B serotype. The serotype identifies the HLA-B*47 gene products. Comparison of B47 nucleotide sequence with other HLA-B sequences shows a segment of 228 bp identical with B44 in the alpha 1 domain and a segment of 218 bp identical with B27 in the alpha 2 domain, but only a 91 bp segment of identity with B13 in the alpha 1 domain. The complex pattern of substitutions and their degree of divergence indicate that HLA-B13 and HLA-Bw47 alleles are not related by a simple mutational event. B47 is linked to a gene that causes adrenal deficiency. B47 is generally low in frequency and with highest known frequencies in Central and Western Africa.
HLA-B46 (B46) is an HLA-B serotype. The serotype identifies the gene products of HLA-B*4601 allele. B*4601 resulted from a rare, interlocus, gene conversion between B62, probably B*1501, and a HLA-C allele. B*4601 is the most common HLA-B allele that does not have an origin within Africa, and estimated 400 million people in Eastern Asia carry a B46 allele. When found B*4601 segregates with only 2 HLA-Cw alleles, A limited number of HLA-A and HLA-DRB1 alleles suggesting that the allele recently expanded from a limited sized group within SE Asia. Extremely low frequencies outside of Eastern Asia are indicators of a recent expansion of B46 from a recently small population. The frequency distribution suggests the ancestral B46 population was in SE China, or, potentially Burma. B46 in Asia correlates with wet-rice farming. The exceptions are notable, it has been found in the Nivkhi on north-eastern Sakalin Island, the Ainu, and the Nivkhi-related (genetically) Tlinglet population of Alaska at trace levels.
HLA-B37 (B37) is an HLA-B serotype. The serotype identifies the more common HLA-B*37 gene products.
HLA-B13 (B13) is an HLA-B serotype. The serotype identifies the more common HLA-B*13 gene products.
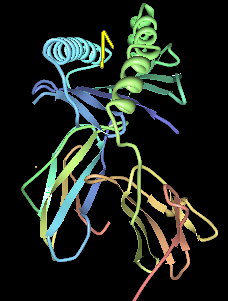
HLA-B8 (B8) is an HLA-B serotype. The serotype identifies the HLA-B*08 gene products. HLA-B8, previously known as HL-A8 was one of the first identified of the HLA antigens. It coined the "Super B8" haplotype, also called the ancestral European haplotype because of its common occurrence in Europe, particular the isles and Scandinavia. B8 is a component gene-allele of the AH8.1 haplotype in Northern and Western Europeans. Genes between B8 and DR3 on this haplotype are frequently associated with autoimmune disease.
HLA-B55 (B55) is an HLA-B serotype. B55 is a split antigen from the B22 broad antigen, sister serotypes are B54 and B56. The serotype identifies the more common HLA-B*55 gene products.
HLA-B54 (B54) is an HLA-B serotype. B54 is a split antigen from the B22 broad antigen, sister serotypes are B55 and B56. The serotype identifies the more common HLA-B*55 gene products.
HLA-B49 (B49) is an HLA-B serotype. B49 is a split antigen from the B21 broad antigen, the sister serotype B50. The serotype identifies the more common HLA-B*50 gene products.
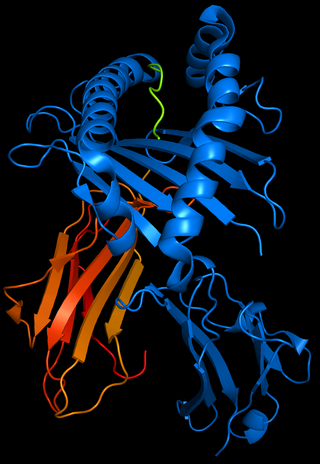
HLA-B51 (B51) is an HLA-B serotype. The serotype identifies the more common HLA-B*51 gene products.
HLA-B39 (B39) is an HLA-B serotype. The serotype identifies the more common HLA-B*39 gene products.
HLA-B38 (B38) is an HLA-B serotype. The serotype identifies the B*38 allele products of the HLA-B gene-locus.
HLA-B70 (B70) is an HLA-B serotype. The serotype identifies certain B*15 gene-allele protein products of HLA-B. B70 is one of many split antigens of the broad antigen, B15.
HLA-B63 (B63) is an HLA-B serotype. The serotype identifies certain B*15 gene-allele protein products of HLA-B.
