Selfish genetic elements are genetic segments that can enhance their own transmission at the expense of other genes in the genome, even if this has no positive or a net negative effect on organismal fitness. Genomes have traditionally been viewed as cohesive units, with genes acting together to improve the fitness of the organism. However, when genes have some control over their own transmission, the rules can change, and so just like all social groups, genomes are vulnerable to selfish behaviour by their parts.
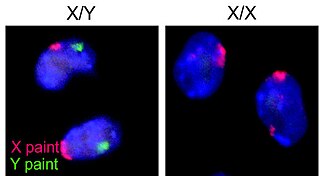
The XY sex-determination system is a sex-determination system used to classify many mammals, including humans, some insects (Drosophila), some snakes, some fish (guppies), and some plants. In this system, the sex of an individual is determined by a pair of sex chromosomes. In most cases, females have two of the same kind of sex chromosome (XX), and are called the homogametic sex. Males have two different kinds of sex chromosomes (XY), and are called the heterogametic sex.

In genetics, dominance is the phenomenon of one variant (allele) of a gene on a chromosome masking or overriding the effect of a different variant of the same gene on the other copy of the chromosome. The first variant is termed dominant and the second is called recessive. This state of having two different variants of the same gene on each chromosome is originally caused by a mutation in one of the genes, either new or inherited. The terms autosomal dominant or autosomal recessive are used to describe gene variants on non-sex chromosomes (autosomes) and their associated traits, while those on sex chromosomes (allosomes) are termed X-linked dominant, X-linked recessive or Y-linked; these have an inheritance and presentation pattern that depends on the sex of both the parent and the child. Since there is only one copy of the Y chromosome, Y-linked traits cannot be dominant or recessive. Additionally, there are other forms of dominance, such as incomplete dominance, in which a gene variant has a partial effect compared to when it is present on both chromosomes and co-dominance, in which different variants on each chromosome both show their associated traits.

Sex linked describes the sex-specific reading patterns of inheritance and presentation when a gene mutation (allele) is present on a sex chromosome (allosome) rather than a non-sex chromosome (autosome). In humans, these are termed X-linked recessive, X-linked dominant and Y-linked. The inheritance and presentation of all three differ depending on the sex of both the parent and the child. This makes them characteristically different from autosomal dominance and recessiveness.

Dosage compensation is the process by which organisms equalize the expression of genes between members of different biological sexes. Across species, different sexes are often characterized by different types and numbers of sex chromosomes. In order to neutralize the large difference in gene dosage produced by differing numbers of sex chromosomes among the sexes, various evolutionary branches have acquired various methods to equalize gene expression among the sexes. Because sex chromosomes contain different numbers of genes, different species of organisms have developed different mechanisms to cope with this inequality. Replicating the actual gene is impossible; thus organisms instead equalize the expression from each gene. For example, in humans, female (XX) cells randomly silence the transcription of one X chromosome, and transcribe all information from the other, expressed X chromosome. Thus, human females have the same number of expressed X-linked genes per cell as do human males (XY), both sexes having essentially one X chromosome per cell, from which to transcribe and express genes.
In biology, outbreeding depression happens when crosses between two genetically distant groups or populations result in a reduction of fitness. The concept is in contrast to inbreeding depression, although the two effects can occur simultaneously. Outbreeding depression is a risk that sometimes limits the potential for genetic rescue or augmentations. It is considered postzygotic response because outbreeding depression is noted usually in the performance of the progeny.
Meiotic drive is a type of intragenomic conflict, whereby one or more loci within a genome will affect a manipulation of the meiotic process in such a way as to favor the transmission of one or more alleles over another, regardless of its phenotypic expression. More simply, meiotic drive is when one copy of a gene is passed on to offspring more than the expected 50% of the time. According to Buckler et al., "Meiotic drive is the subversion of meiosis so that particular genes are preferentially transmitted to the progeny. Meiotic drive generally causes the preferential segregation of small regions of the genome".
The mechanisms of reproductive isolation are a collection of evolutionary mechanisms, behaviors and physiological processes critical for speciation. They prevent members of different species from producing offspring, or ensure that any offspring are sterile. These barriers maintain the integrity of a species by reducing gene flow between related species.

The heterogametic sex is the sex of a species where an individual's gametes have non-matching sex chromosomes. In humans, the heterogametic sex is the male sex, where each gamete's sex chromosomes are X and Y. This is in contrast to the female sex, where each gamete's sex chromosomes are X and X. This arrangement is understood within the XY sex-determination system.
Sex chromosomes are chromosomes that carry the genes that determine the sex of an individual. The human sex chromosomes are a typical pair of mammal allosomes. They differ from autosomes in form, size, and behavior. Whereas autosomes occur in homologous pairs whose members have the same form in a diploid cell, members of an allosome pair may differ from one another.
Hybrid growth disorders refer to reduced growth or overgrowth in an organism that is a hybrid of two different species. In some sense, it is a type of hybrid dysgenesis when the growth disorder proves deleterious, making it the opposite of heterosis or hybrid vigour.
H. Allen Orr is the Shirley Cox Kearns Professor of Biology at the University of Rochester.
The concept of a biological species as a group of organisms capable of interbreeding to produce viable offspring dates back to at least the 18th century, although it is often associated today with Ernst Mayr. Species of the fruit-fly Drosophila are one of the most commonly used organisms in evolutionary research, and have been used to test many theories related to the evolution of species. The genus Drosophila comprises numerous species that have varying degrees of premating and postmating isolation between them. These species are useful for testing hypotheses of the reproductive mechanisms underlying speciation.

The Bateson–Dobzhansky–Muller model, also known as Dobzhansky–Muller model, is a model of the evolution of genetic incompatibility, important in understanding the evolution of reproductive isolation during speciation and the role of natural selection in bringing it about. The theory was first described by William Bateson in 1909, then independently described by Theodosius Dobzhansky in 1934, and later elaborated in different forms by Herman Muller, H. Allen Orr and Sergey Gavrilets.

Reinforcement is a process of speciation where natural selection increases the reproductive isolation between two populations of species. This occurs as a result of selection acting against the production of hybrid individuals of low fitness. The idea was originally developed by Alfred Russel Wallace and is sometimes referred to as the Wallace effect. The modern concept of reinforcement originates from Theodosius Dobzhansky. He envisioned a species separated allopatrically, where during secondary contact the two populations mate, producing hybrids with lower fitness. Natural selection results from the hybrid's inability to produce viable offspring; thus members of one species who do not mate with members of the other have greater reproductive success. This favors the evolution of greater prezygotic isolation. Reinforcement is one of the few cases in which selection can favor an increase in prezygotic isolation, influencing the process of speciation directly. This aspect has been particularly appealing among evolutionary biologists.
Caenorhabditis nigoni is a male-female species in the Elegans group of the genus Caenorhabditis, first identified and described as "Caenorhabditis species 9" or "C. sp. 9" before being renamed as "C. nigoni". The specific epithet is a tribute to Victor Nigon who first studied Caenorhabditis elegans in the laboratory with Ellsworth Dougherty in the 1940s. Isolates come from the Democratic Republic of the Congo and Kerala, India.
This glossary of genetics and evolutionary biology is a list of definitions of terms and concepts used in the study of genetics and evolutionary biology, as well as sub-disciplines and related fields, with an emphasis on classical genetics, quantitative genetics, population biology, phylogenetics, speciation, and systematics. Overlapping and related terms can be found in Glossary of cellular and molecular biology, Glossary of ecology, and Glossary of biology.
Hybrid incompatibility is a phenomenon in plants and animals, wherein offspring produced by the mating of two different species or populations have reduced viability and/or are less able to reproduce. Examples of hybrids include mules and ligers from the animal world, and subspecies of the Asian rice crop Oryza sativa from the plant world. Multiple models have been developed to explain this phenomenon. Recent research suggests that the source of this incompatibility is largely genetic, as combinations of genes and alleles prove lethal to the hybrid organism. Incompatibility is not solely influenced by genetics, however, and can be affected by environmental factors such as temperature. The genetic underpinnings of hybrid incompatibility may provide insight into factors responsible for evolutionary divergence between species.
Eukaryote hybrid genomes result from interspecific hybridization, where closely related species mate and produce offspring with admixed genomes. The advent of large-scale genomic sequencing has shown that hybridization is common, and that it may represent an important source of novel variation. Although most interspecific hybrids are sterile or less fit than their parents, some may survive and reproduce, enabling the transfer of adaptive variants across the species boundary, and even result in the formation of novel evolutionary lineages. There are two main variants of hybrid species genomes: allopolyploid, which have one full chromosome set from each parent species, and homoploid, which are a mosaic of the parent species genomes with no increase in chromosome number.
Genetic incompatibility describes the process by which mating yields offspring that are nonviable, prone to disease, or genetically defective in some way. In nature, animals can ill afford to devote costly resources for little or no reward, ergo, mating strategies have evolved to allow females to choose or otherwise determine mates which are more likely to result in viable offspring.

