| "Parvarchaeota" | |
|---|---|
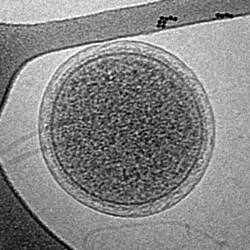 | |
| Parvarchaeum acidiphilum | |
| Scientific classification | |
| Domain: | Archaea |
| Kingdom: | Nanobdellati |
| Phylum: | "Parvarchaeota" Rinke et al. 2013 |
| Orders | |
| |
"Parvarchaeota" is a phylum of archaea belonging to the Nanobdellati kingdom. They have been discovered in acid mine drainage waters and later in marine sediments. The cells of these organisms are extremely small consistent with small genomes. Metagenomic techniques allow obtaining genomic sequences from non-cultured organisms, which were applied to determine this phylum. [1]
Contents
The type species is "Candidatus Parvarchaeum acidiphilum". They have very small cells, around 400-500 nm, and reduced genomes made up of about 1000 genes. A similar-sized archaea that has been found in the same acidic environments is "Ca. Microarcheum", from the phylum Micrarchaeota. [1] [2]
According to the phylogenetic trees "Parvarchaeota" may be a sister group of Thermoplasmata within Euryarchaeota [3] or belong to Nanobdellati, [1] although it has also been suggested that all the Nanobdellati archaea belong phylogenetically to Euryarchaeota. [3] [4]