
A chordate is a deuterostomic animal belonging to the phylum Chordata. All chordates possess, at some point during their larval or adult stages, five distinctive physical characteristics (synapomorphies) that distinguish them from other taxa. These five synapomorphies are a notochord, a hollow dorsal nerve cord, an endostyle or thyroid, pharyngeal slits, and a post-anal tail. The name "chordate" comes from the first of these synapomorphies, the notochord, which plays a significant role in chordate body plan structuring and movements. Chordates are also bilaterally symmetric, have a coelom, possess a closed circulatory system, and exhibit metameric segmentation.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.
Lethiscus is the earliest known representative of the Aistopoda, a group of very specialised snake-like tetrapodomorphs known from the early Carboniferous (Mississippian).

Temnospondyli or temnospondyls is a diverse ancient order of small to giant tetrapods—often considered primitive amphibians—that flourished worldwide during the Carboniferous, Permian and Triassic periods, with fossils being found on every continent. A few species continued into the Jurassic and Early Cretaceous periods, but all had gone extinct by the Late Cretaceous. During about 210 million years of evolutionary history, they adapted to a wide range of habitats, including freshwater, terrestrial, and even coastal marine environments. Their life history is well understood, with fossils known from the larval stage, metamorphosis and maturity. Most temnospondyls were semiaquatic, although some were almost fully terrestrial, returning to the water only to breed. These temnospondyls were some of the first vertebrates fully adapted to life on land. Although temnospondyls are amphibians, many had characteristics such as scales and armour-like bony plates that distinguish them from the modern soft-bodied lissamphibians.

Suminia is an extinct genus of basal anomodont that lived during the Tatarian age of the late Permian, spanning approximately from 268-252 Ma. Suminia is recognized the youngest non-dicynodont anomodont. Its fossil localities are primarily derived from the Kotel’nich locality of the Kirov Oblast in Russia. However, there have been some isolated specimen found in a few different localities, all from eastern European regions of Russia.

Westlothiana is a genus of reptile-like tetrapod that lived about 338 million years ago during the latest part of the Viséan age of the Carboniferous. Members of the genus bore a superficial resemblance to modern-day lizards. The genus is known from a single species, Westlothiana lizziae. The type specimen was discovered in the East Kirkton Limestone at the East Kirkton Quarry, West Lothian, Scotland in 1984. This specimen was nicknamed "Lizzie the lizard" by fossil hunter Stan Wood, and this name was quickly adopted by other paleontologists and the press. When the specimen was formally named in 1990, it was given the specific name "lizziae" in homage to this nickname. However, despite its similar body shape, Westlothiana is not considered a true lizard. Westlothiana's anatomy contained a mixture of both "labyrinthodont" and reptilian features, and was originally regarded as the oldest known reptile or amniote. However, updated studies have shown that this identification is not entirely accurate. Instead of being one of the first amniotes, Westlothiana was rather a close relative of Amniota. As a result, most paleontologists since the original description place the genus within the group Reptiliomorpha, among other amniote relatives such as diadectomorphs and seymouriamorphs. Later analyses usually place the genus as the earliest diverging member of Lepospondyli, a collection of unusual tetrapods which may be close to amniotes or lissamphibians, or potentially both at the same time.

Nectridea is the name of an extinct order of lepospondyl tetrapods from the Carboniferous and Permian periods, including animals such as Diplocaulus. In appearance, they would have resembled modern newts or aquatic salamanders, although they are not close relatives of modern amphibians. They were characterized by long, flattened tails to aid in swimming, as well as numerous features of the vertebrae.

Metoposaurus meaning "front lizard" is an extinct genus of stereospondyl temnospondyl amphibian, known from the Late Triassic of Germany, Italy, Poland, and Portugal. This mostly aquatic animal possessed small, weak limbs, sharp teeth, and a large, flat head. This highly flattened creature mainly fed on fish, which it captured with its wide jaws lined with needle-like teeth. Many Metoposaurus mass graves have been found, probably from creatures that grouped together in drying pools during drought.

Embolomeri is an order of tetrapods or stem-tetrapods, possibly members of Reptiliomorpha. Embolomeres first evolved in the Early Carboniferous (Mississippian) Period and were the largest and most successful predatory tetrapods of the Late Carboniferous (Pennsylvanian) Period. They were specialized semiaquatic predators with long bodies for eel-like undulatory swimming. Embolomeres are characterized by their vertebral centra, which are formed by two cylindrical segments, the pleurocentrum at the rear and intercentrum at the front. These segments are equal in size. Most other tetrapods have pleurocentra and intercentra which are drastically different in size and shape.

Branchiosaurus is a genus of small, prehistoric amphibians. Fossils have been discovered in strata dating from the late Pennsylvanian Epoch to the Permian Period. The taxa may be invalid; the material referred to the genus may be juvenile specimens of larger amphibians.
Pachycheilosuchus is an extinct genus of neosuchian from the Early Cretaceous of Texas, United States. Previously known, in part, as the "Glen Rose form", this crocodylomorph is notable for its procoelous vertebrae, otherwise found only in derived eusuchian crocodilians, a thick margin on the maxillae, and a shield of armor on the neck formed by the fusion of six individual scutes.

Acherontiscus is an extinct genus of stegocephalians that lived in the Early Carboniferous of Scotland. The type and only species is Acherontiscus caledoniae, named by paleontologist Robert Carroll in 1969. Members of this genus have an unusual combination of features which makes their placement within amphibian-grade tetrapods uncertain. They possess multi-bone vertebrae similar to those of embolomeres, but also a skull similar to lepospondyls. The only known specimen of Acherontiscus possessed an elongated body similar to that of a snake or eel. No limbs were preserved, and evidence for their presence in close relatives of Acherontiscus is dubious at best. Phylogenetic analyses created by Marcello Ruta and other paleontologists in the 2000s indicate that Acherontiscus is part of Adelospondyli, closely related to other snake-like animals such as Adelogyrinus and Dolichopareias. Adelospondyls are traditionally placed within the group Lepospondyli due to their fused vertebrae. Some analyses published since 2007 have argued that adelospondyls such as Acherontiscus may not actually be lepospondyls, instead being close relatives or members of the family Colosteidae. This would indicate that they evolved prior to the split between the tetrapod lineage that leads to reptiles (Reptiliomorpha) and the one that leads to modern amphibians (Batrachomorpha). Members of this genus were probably aquatic animals that were able to swim using snake-like movements.
Kourerpeton is an extinct genus of dvinosaurian temnospondyl. Fossils of Kourerpeton were discovered in a window of a barber's shop in either Bisbee or Mesa, Arizona. Kourerpeton was named in 1976, with the type and only species being K. bradyi. It was originally assigned to the monotypic family Kourerpetidae, which has been alternatively spelled Kourerpetontidae.
Trihecaton is an extinct genus of microsaur from the Late Pennsylvanian of Colorado. Known from a single species, Trihecaton howardinus, this genus is distinctive compared to other microsaurs due to possessing a number of plesiomorphic ("primitive") features relative to the rest of the group. These include large intercentra, folded enamel, and a large coronoid process of the jaw. Its classification is controversial due to combining a long body with strong limbs, features which typically are not present at the same time in other microsaurs. Due to its distinctiveness, Trihecaton has been given its own monospecific family, Trihecatontidae.
Slaugenhopia is an extinct genus of dvinosaurian temnospondyl within the family Tupilakosauridae. Fossils have been found from the Early Permian San Angelo Formation in Texas. The type and only species, S. texensis, was named in 1962. It may be closely related to the dvinosaur Kourerpeton. Slaugenhopia was once classified as a trimerorhachid but is now classified as a tupilakosaurid.
Antlerpeton is an extinct genus of early tetrapod from the Early Carboniferous of Nevada. It is known from a single poorly preserved skeleton from the Diamond Peak Formation of Eureka County. A mix of features in its compound vertebrae suggest that Antlerpeton is a primitive stem tetrapod that has affinities with later, more advanced forms. Its robust pelvis and hind limbs allowed for effective locomotion on land, but the animal was likely still tied to a semiaquatic lifestyle near the coast.

The vertebral column, also known as the backbone or spine, is the core part of the axial skeleton in vertebrate animals. The vertebral column is the defining characteristic of vertebrate endoskeleton in which the notochord found in all chordates has been replaced by a segmented series of mineralized irregular bones called vertebrae, separated by fibrocartilaginous intervertebral discs. The dorsal portion of the vertebral column houses the spinal canal, a cavity formed by alignment of the neural arches that encloses and protects the spinal cord.

Albertonectes is an extinct genus of elasmosaurid plesiosaur known from the Late Cretaceous Bearpaw Formation of Alberta, Canada. It contains a single species, Albertonectes vanderveldei. Albertonectes is the longest elasmosaur, and more generally plesiosaur, known to date both in neck and total body length.

Rhachitomi is a group of temnospondyl amphibians that includes all temnospondyls except edopoids and dendrerpetontids. It was established as a clade name by German paleontologist Rainer R. Schoch in 2013, although the name had first been established in 1919 by British paleontologist D. M. S. Watson to encompass an evolutionary grade of temnospondyls leading to the group Stereospondyli. American paleontologist Alfred Romer used the term in a similar sense, grouping most Permian and Triassic temnospondyls under Rhachitomi. A similar name that appeared earlier in the scientific literature is Rachitomi, which was named by American paleontologist Edward Drinker Cope in 1882. Rachitomi was commonly used in the late nineteenth and early twentieth century to include early amphibians such as Eryops and Archegosaurus that had rhachitomous vertebrae. Many early tetrapods have vertebrae that are split into two parts below the notochord: a pleurocentrum and an intercentrum. In rhachitomous vertebrae, the intercentrum is large and semicircular, while the pleurocentrum divided into two smaller paired elements. Schoch defined Rhachitomi as a node-based taxon to include four major and well-supported clades of temnospondyls: Dvinosauria, Eryopidae, Stereospondyli and a clade formed by Zatracheidae and Dissorophoidea. Not all members of Rhachitomi have rhachitomous vertebrae; the largest subgroup, Stereospondyli, lacks pleurocentra. Below is a cladogram from Schoch's analysis showing the placement of Rhachitomi within Temnospondyli:
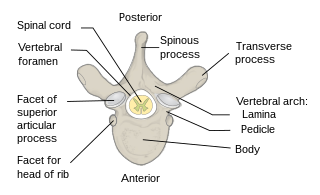
Each vertebra is an irregular bone with a complex structure composed of bone and some hyaline cartilage, that make up the vertebral column or spine, of vertebrates. The proportions of the vertebrae differ according to their spinal segment and the particular species.







