
Ophidia is a group of squamate reptiles including modern snakes and reptiles more closely related to snakes than to other living groups of lizards.

Huabeisaurus was a genus of dinosaur from the Late Cretaceous. It was a sauropod which lived in what is present-day northern China. The type species, Huabeisaurus allocotus, was first described by Pang Qiqing and Cheng Zhengwu in 2000. Huabeisaurus is known from numerous remains found in the 1990s, which include teeth, partial limbs and vertebrae. Due to its relative completeness, Huabeisaurus represents a significant taxon for understanding sauropod evolution in Asia. Huabeisaurus comes from Kangdailiang and Houyu, Zhaojiagou Town, Tianzhen County, Shanxi province, China. The holotype was found in the unnamed upper member of the Huiquanpu Formation, which is Late Cretaceous (?Cenomanian–?Campanian) in age based on ostracods, charophytes, and fission-track dating.

Pythonomorpha was originally proposed by paleontologist Edward Drinker Cope (1869) as a reptilian order comprising mosasaurs, which he believed to be close relatives of Ophidia (snakes). The etymology of the term Pythonomorpha comes from the Greek Python and morphe ("form"), and refers to the generally serpentine body plan of members of the group. Cope wrote, "In the mosasauroids, we almost realize the fictions of snake-like dragons and sea-serpents, in which men have been ever prone to indulge. On account of the ophidian part of their affinities, I have called this order Pythonomorpha." Cope incorporated two families, the Clidastidae and the Mosasauridae.

Nanshiungosaurus is a genus of therizinosaurid that lived in what is now Asia during the Late Cretaceous of South China. The type species, Nanshiungosaurus brevispinus, was first discovered in 1974 and described in 1979 by Dong Zhiming. It is represented by a single specimen preserving most of the cervical and dorsal vertebrae with the pelvis. A supposed and unlikely second species, "Nanshiungosaurus" bohlini, was found in 1992 and described in 1997. It is also represented by vertebrae but this species however, differs in geological age and lacks authentic characteristics compared to the type, making its affinity to the genus unsupported.
Muraenosaurus is an extinct genus of cryptoclidid plesiosaur reptile from the Oxford Clay of Southern England. The genus was given its name due to the eel-like appearance of the long neck and small head. Muraenosaurus grew up to 5.2 metres (17 ft) in length and lived roughly between 160 Ma and 164 Ma in the Callovian of the middle Jurassic. Charles E. Leeds collected the first Muraenosaurus which was then described by H. G. Seeley. The specimen may have suffered some damage due to the casual style of Charles Leeds’ collection. The first muraenosaur was recovered with pieces missing from the skull and many of the caudal vertebrae absent. Because the animal was described from Charles Leeds’ collection it was given the name Muraenosaurus Leedsi. M. leedsi is the most complete specimen belonging to the genus Muraenosaurus and also the only species that is undoubtedly a member of the genus. Two other species have been tentatively referred to as members of the genus Muraenosaurus: M. reedii and Muraenosaurus beloclis Seeley 1892, which in 1909 became the separate genus Picrocleidus.
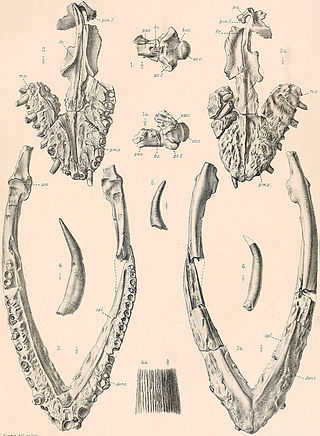
Platecarpus is an extinct genus of aquatic lizards belonging to the mosasaur family, living around 84–81 million years ago during the middle Santonian to early Campanian, of the Late Cretaceous period. Fossils have been found in the United States and possible specimens in Belgium and Africa. A well-preserved specimen of Platecarpus shows that it fed on moderate-sized fish, and it has been hypothesized to have fed on squid, and ammonites as well. Like other mosasaurs, it was initially thought to have swum in an eel-like fashion, although another study suggests that it swam more like modern sharks. An exceptionally well-preserved specimen of P. tympaniticus known as LACM 128319 shows skin impressions, pigments around the nostrils, bronchial tubes, and the presence of a high-profile tail fluke, showing that it and other mosasaurs did not necessarily have an eel-like swimming method, but were more powerful, fast swimmers. It is held in the Natural History Museum of Los Angeles County. Isotopic analysis on teeth specimens has suggested that this genus and Clidastes may have entered freshwater occasionally, just like modern sea snakes.

Kaganaias is an extinct genus of basal and oldest dolichosaur that lived in what is now Japan during the Early Cretaceous. Kaganaias was semi-aquatic and is the only known aquatic squamate known from before the Cenomanian stage of the Cretaceous. It is also the first to be found in an inland area, instead of on the coast where aquatic squamates are commonly found. Its generic name is derived from Kaga Province, the old name for the Ishikawa Prefecture where the specimens were found, while the species name hakusanensis comes from the mountain that gives its name to Hakusan the city near its find site. The geological formation in which the specimens were found, the Kuwajima Formation, stands alongside the Tetori River and has been the site of numerous other finds including molluscs, dinosaurs, fish, and pterosaurs.

Mahajangasuchus is an extinct genus of crocodyliform which had blunt, conical teeth. The type species, M. insignis, lived during the Late Cretaceous; its fossils have been found in the Maevarano Formation in northern Madagascar. It was a fairly large predator, measuring up to 4 metres (13 ft) long.

Taniwhasaurus is an extinct genus of mosasaurs that lived during the Campanian stage of the Late Cretaceous. It is a member of the subfamily Tylosaurinae, a lineage of mosasaurs characterized by a long toothless conical rostrum. Two valid species are attached to the genus, T. oweni and T. antarcticus, known respectively from the fossil record of present-day New Zealand and Antarctica. Two other species have been nominally classified within the genus, T. 'capensis' and T. 'mikasaensis', recorded in present-day South Africa and Japan, but their attribution remains problematic due to the fragmentary state of their fossils. The generic name literally means "taniwha lizard", referring to a supernatural aquatic creature from Māori mythology.

Eupodophis is an extinct genus of snake from the Late Cretaceous period. It has two small hind legs and is considered a transitional form between Cretaceous lizards and limbless snakes. The feature, described as vestigial, was most likely useless to Eupodophis. The type species Eupodophis descouensi was named in 2000 and resides now in the paleontology section of the Mim Museum in Beirut, Lebanon. The specific name is dedicated to the French naturalist Didier Descouens.

Tatenectes is a genus of cryptoclidid plesiosaur known from the Upper Jurassic of Wyoming. Its remains were recovered from the Redwater Shale Member of the Sundance Formation, and initially described as a new species of Cimoliosaurus by Wilbur Clinton Knight in 1900. It was reassigned to Tricleidus by Maurice G. Mehl in 1912 before being given its own genus by O'Keefe and Wahl in 2003. Tatenectes laramiensis is the type and only species of Tatenectes. While the original specimen was lost, subsequent discoveries have revealed that Tatenectes was a very unusual plesiosaur. Its torso had a flattened, boxy cross-section and its gastralia exhibit pachyostosis (thickening). The total length of Tatenectes has been estimated at 2–3 meters (6.6–9.8 ft).

Carsosaurus is a genus of extinct amphibious reptiles, in the mosasaur superfamily, containing only the species Carsosaurus marchesetti. It is known from a single individual that lived during the Upper Cretaceous in what is now Slovenia. The specimen is well-preserved, containing many different bones as well as some skin impressions and sternal cartilage. While more remains are needed to be certain, it is generally thought to belong to the Aigialosauridae. In life, it was an amphibious creature that spent most of its time on land, although its later relatives would become fully aquatic.
Parviraptor is a genus of squamate containing one species, Parviraptor estesi, from the Late Jurassic (Tithonian) or Early Cretaceous (Berriasian) Purbeck Limestone Formation of Dorset, England. A second species, Parviraptor gilmorei, was described from the Late Jurassic Morrison Formation of Western North America; it was present in stratigraphic zone 4. However, the second species was subsequently transferred to a separate genus Diablophis. An indeterminate species is known from the Bathonian aged Kirtlington Mammal Bed.
Acteosaurus is an extinct genus of aquatic lizard that lived in the upper Cretaceous period. Its species, A. tommasinii and A. crassicostatus, were described in 1860 and 1993. Though A. crassicostatus is probably a junior synonym for Adriosaurus suessi, A. tommasinii was found to be similar to coniasaurs, mosasauroids, and a sister taxon to modern snakes in 2010.
Eremiasaurus is a genus of mosasaurs, an extinct group of marine reptiles. It lived during the Maastrichtian stage of the Late Cretaceous in what is now North Africa. Only one species is known, E. heterodontus, described in 2012 from two remarkably complete fossil specimens discovered in the Ouled Abdoun Basin, Morocco. This site is known to have delivered a significant number of other related mosasaurs.

The Tethysaurinae are a subfamily of mosasaurs, a diverse group of Late Cretaceous marine squamates. Members of the subfamily are informally and collectively known as "tethysaurines" and have been recovered from North America and Africa. Only two tethysaurine genera are known, Pannoniasaurus and Tethysaurus. The genera Yaguarasaurus and Russellosaurus were previously considered tethysaurines until they were grouped with Romeosaurus in the new subfamily Yaguarasaurinae. A possible member of this clade (subfamily) is a mosasaur specimen known from a maxilla fragment, found in 1960 in the Czech Republic, in Dolní Újezd near Litomyšl.

The Yaguarasaurinae are a subfamily of mosasaurs, a diverse group of Late Cretaceous marine squamates. Members of the subfamily are informally and collectively known as "yaguarasaurines" and have been recovered from North and South America and Europe. Three genera, Yaguarasaurus, Russellosaurus and Romeosaurus are known. Yaguarasaurus and Russellosaurus were previously considered part of the Tethysaurinae until they were grouped with Romeosaurus as yaguarasaurines.

Tetrapodophis is an extinct genus of lizard from the Early Cretaceous (Aptian) aged Crato Formation of Brazil. It has an elongate snake-like body, with four limbs. Tetrapodophis has been considered by some authors to be one of the oldest members of Ophidia. However, this classification has been disputed by some other authors, who identify Tetrapodophis as a dolichosaurid much less closely related to snakes.

Pachyophis is an extinct genus of Simoliophiidae snakes that were extant during the Cenomanian stage of the Late Cretaceous period. More specifically, it was found to be from the Cenomanian Age about 93.9-100.5 million years ago in the suburb area of Bileca, Herzegovina.

Dolichosauridae is a family of Cretaceous aquatic lizards. They are widely considered to be the earliest and most primitive members of Mosasauria, though some researchers have recovered them as more closely related to snakes.

