Glycoproteins are proteins which contain oligosaccharide (sugar) chains covalently attached to amino acid side-chains. The carbohydrate is attached to the protein in a cotranslational or posttranslational modification. This process is known as glycosylation. Secreted extracellular proteins are often glycosylated.

Lipopolysaccharide, now more commonly known as endotoxin, is a collective term for components of the outermost membrane of cell envelope of gram-negative bacteria, such as E. coli and Salmonella with a common structural architecture. Lipopolysaccharides (LPS) are large molecules consisting of three parts: an outer core polysaccharide termed the O-antigen, an inner core oligosaccharide and Lipid A, all covalently linked. In current terminology, the term endotoxin is often used synonymously with LPS, although there are a few endotoxins that are not related to LPS, such as the so-called delta endotoxin proteins produced by Bacillus thuringiensis.
An oligosaccharide is a saccharide polymer containing a small number of monosaccharides. Oligosaccharides can have many functions including cell recognition and cell adhesion.

The plasma membranes of cells contain combinations of glycosphingolipids, cholesterol and protein receptors organised in glycolipoprotein lipid microdomains termed lipid rafts. Their existence in cellular membranes remains controversial. Indeed, Kervin and Overduin imply that lipid rafts are misconstrued protein islands, which they propose form through a proteolipid code. Nonetheless, it has been proposed that they are specialized membrane microdomains which compartmentalize cellular processes by serving as organising centers for the assembly of signaling molecules, allowing a closer interaction of protein receptors and their effectors to promote kinetically favorable interactions necessary for the signal transduction. Lipid rafts influence membrane fluidity and membrane protein trafficking, thereby regulating neurotransmission and receptor trafficking. Lipid rafts are more ordered and tightly packed than the surrounding bilayer, but float freely within the membrane bilayer. Although more common in the cell membrane, lipid rafts have also been reported in other parts of the cell, such as the Golgi apparatus and lysosomes.
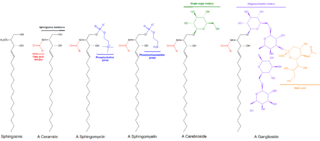
Sphingolipids are a class of lipids containing a backbone of sphingoid bases, which are a set of aliphatic amino alcohols that includes sphingosine. They were discovered in brain extracts in the 1870s and were named after the mythological sphinx because of their enigmatic nature. These compounds play important roles in signal transduction and cell recognition. Sphingolipidoses, or disorders of sphingolipid metabolism, have particular impact on neural tissue. A sphingolipid with a terminal hydroxyl group is a ceramide. Other common groups bonded to the terminal oxygen atom include phosphocholine, yielding a sphingomyelin, and various sugar monomers or dimers, yielding cerebrosides and globosides, respectively. Cerebrosides and globosides are collectively known as glycosphingolipids.
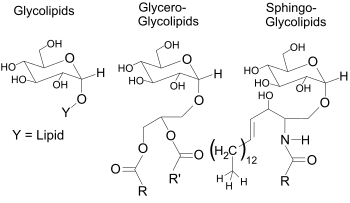
Glycosphingolipids are a subtype of glycolipids containing the amino alcohol sphingosine. They may be considered as sphingolipids with an attached carbohydrate. Glycosphingolipids are a group of lipids and are a part of the cell membrane. They consist of a hydrophobic ceramide part and a glycosidically bound carbohydrate part. This oligosaccharide content remains on the outside of the cell membrane where it is important for biological processes such as cell adhesion or cell–cell interactions. Glycosphingolipids play also important role in oncogenesis and ontogenesis.

A ganglioside is a molecule composed of a glycosphingolipid with one or more sialic acids linked on the sugar chain. NeuNAc, an acetylated derivative of the carbohydrate sialic acid, makes the head groups of gangliosides anionic at pH 7, which distinguishes them from globosides.
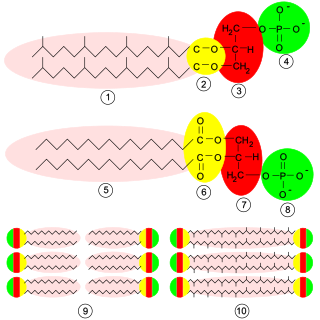
Glycerophospholipids or phosphoglycerides are glycerol-based phospholipids. They are the main component of biological membranes in eukaryotic cells. They are a type of lipid, of which its composition affects membrane structure and properties. Two major classes are known: those for bacteria and eukaryotes and a separate family for archaea.
CD1 is a family of glycoproteins expressed on the surface of various human antigen-presenting cells. CD1 glycoproteins are structurally related to the class I MHC molecules, however, in contrast to MHC class 1 proteins, they present lipids, glycolipids and small molecules antigens, from both endogenous and pathogenic proteins, to T cells and activate an immune response. Both αβ and γδ T cells recognise CD1 molecules.
Cerebrosides (monoglycosylceramides) are a group of glycosphingolipids which are important components of animal muscle and nerve cell membranes.
Sulfatide, also known as 3-O-sulfogalactosylceramide, SM4, or sulfated galactocerebroside, is a class of sulfolipids, specifically a class of sulfoglycolipids, which are glycolipids that contain a sulfate group. Sulfatide is synthesized primarily starting in the endoplasmic reticulum and ending in the Golgi apparatus where ceramide is converted to galactocerebroside and later sulfated to make sulfatide. Of all of the galactolipids that are found in the myelin sheath, one fifth of them are sulfatide. Sulfatide is primarily found on the extracellular leaflet of the myelin plasma membrane produced by the oligodendrocytes in the central nervous system and in the Schwann cells in the peripheral nervous system. However, sulfatide is also present on the extracellular leaflet of the plasma membrane of many cells in eukaryotic organisms.
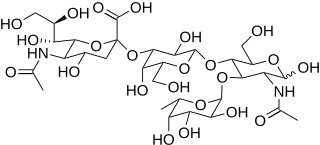
Sialyl LewisX (sLeX), also known as cluster of differentiation 15s (CD15s) or stage-specific embryonic antigen 1 (SSEA-1), is a tetrasaccharide carbohydrate which is usually attached to O-glycans on the surface of cells. It is known to play a vital role in cell-to-cell recognition processes. It is also the means by which an egg attracts sperm; first, to stick to it, then bond with it and eventually form a zygote.
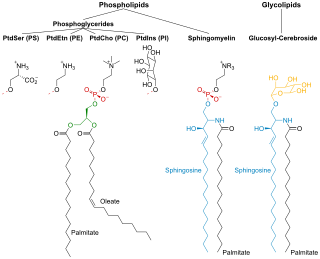
Membrane lipids are a group of compounds which form the lipid bilayer of the cell membrane. The three major classes of membrane lipids are phospholipids, glycolipids, and cholesterol. Lipids are amphiphilic: they have one end that is soluble in water ('polar') and an ending that is soluble in fat ('nonpolar'). By forming a double layer with the polar ends pointing outwards and the nonpolar ends pointing inwards membrane lipids can form a 'lipid bilayer' which keeps the watery interior of the cell separate from the watery exterior. The arrangements of lipids and various proteins, acting as receptors and channel pores in the membrane, control the entry and exit of other molecules and ions as part of the cell's metabolism. In order to perform physiological functions, membrane proteins are facilitated to rotate and diffuse laterally in two dimensional expanse of lipid bilayer by the presence of a shell of lipids closely attached to protein surface, called annular lipid shell.
Carbohydrate–protein interactions are the intermolecular and intramolecular interactions between protein and carbohydrate moieties. These interactions form the basis of specific recognition of carbohydrates by lectins. Carbohydrates are important biopolymers and have a variety of functions. Often carbohydrates serve a function as a recognition element. That is, they are specifically recognized by other biomolecules. Proteins which bind carbohydrate structures are known as lectins. Compared to the study of protein–protein and protein–DNA interaction, it is relatively recent that scientists get to know the protein–carbohydrate binding.
A kodecyte (ko•de•cyte) is a living cell that has been modified (koded) by the incorporation of one or more function-spacer-lipid constructs to gain a new or novel biological, chemical or technological function. The cell is modified by the lipid tail of the FSL construct incorporating into the bilipid membrane of the cell.

In cellular biology, cell–cell recognition is a cell's ability to distinguish one type of neighboring cell from another. This phenomenon occurs when complementary molecules on opposing cell surfaces meet. A receptor on one cell surface binds to its specific ligand on a nearby cell, initiating a cascade of events which regulate cell behaviors ranging from simple adhesion to complex cellular differentiation. Like other cellular functions, cell-cell recognition is impacted by detrimental mutations in the genes and proteins involved and is subject to error. The biological events that unfold due to cell-cell recognition are important for animal development, microbiomes, and human medicine.
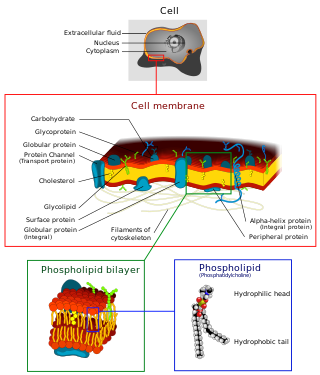
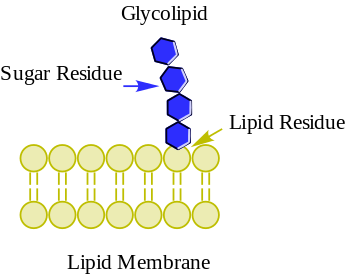
The cell membrane is a biological membrane that separates and protects the interior of a cell from the outside environment. The cell membrane consists of a lipid bilayer, made up of two layers of phospholipids with cholesterols interspersed between them, maintaining appropriate membrane fluidity at various temperatures. The membrane also contains membrane proteins, including integral proteins that span the membrane and serve as membrane transporters, and peripheral proteins that loosely attach to the outer (peripheral) side of the cell membrane, acting as enzymes to facilitate interaction with the cell's environment. Glycolipids embedded in the outer lipid layer serve a similar purpose.
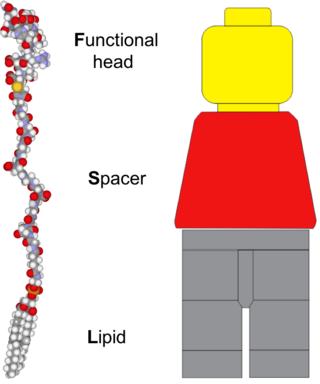
Function-Spacer-Lipid (FSL) Kode constructs are amphiphatic, water dispersible biosurface engineering constructs that can be used to engineer the surface of cells, viruses and organisms, or to modify solutions and non-biological surfaces with bioactives. FSL Kode constructs spontaneously and stably incorporate into cell membranes. FSL Kode constructs with all these aforementioned features are also known as Kode Constructs. The process of modifying surfaces with FSL Kode constructs is known as "koding" and the resultant "koded" cells, viruses and liposomes are respectively known as kodecytes, and kodevirions.

Milk fat globule membrane (MFGM) is a complex and unique structure composed primarily of lipids and proteins that surrounds milk fat globule secreted from the milk producing cells of humans and other mammals. It is a source of multiple bioactive compounds, including phospholipids, glycolipids, glycoproteins, and carbohydrates that have important functional roles within the brain and gut.

T-cell surface glycoprotein CD1b is a protein that in humans is encoded by the CD1B gene.