
A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

Acrophialophora fusispora is a poorly studied ascomycete fungus found in soil, air and various plants. A. fusispora is morphologically similar to the genera Paecilomyces and Masonia, but differ in the presence of pigmented conidiophores, verticillate phialides, and frequent sympodial proliferation. Moreover, A. fusispora is distinguished by its pigmented spindle-shaped conidia, covered with spiral bands. The fungus is naturally found in soils of tropical to temperate regions. The fungus has been identified as a plant and animal pathogen, and has recently been recognized as an emerging opportunistic human pathogen. A. fusispora infection in human is rare and has few documented clinical cases, but due to the rarity of the fungus and potential misidentification, the infections may be underdiagnosed. Clinical cases of A. fusispora include cases of keratitis, pulmonary colonization and infection, and cerebral infections. The fungus also has two documented cases of infection in dogs.

Purpureocillium lilacinum is a species of filamentous fungus in the family Ophiocordycipitaceae. It has been isolated from a wide range of habitats, including cultivated and uncultivated soils, forests, grassland, deserts, estuarine sediments and sewage sludge, and insects. It has also been found in nematode eggs, and occasionally from females of root-knot and cyst nematodes. In addition, it has frequently been detected in the rhizosphere of many crops. The species can grow at a wide range of temperatures – from 8 to 38 °C for a few isolates, with optimal growth in the range 26 to 30 °C. It also has a wide pH tolerance and can grow on a variety of substrates. P. lilacinum has shown promising results for use as a biocontrol agent to control the growth of destructive root-knot nematodes.
Aspergillus ochraceus is a mold species in the genus Aspergillus known to produce the toxin ochratoxin A, one of the most abundant food-contaminating mycotoxins, and citrinin. It also produces the dihydroisocoumarin mellein. It is a filamentous fungus in nature and has characteristic biseriate conidiophores. Traditionally a soil fungus, has now began to adapt to varied ecological niches, like agricultural commodities, farmed animal and marine species. In humans and animals the consumption of this fungus produces chronic neurotoxic, immunosuppressive, genotoxic, carcinogenic and teratogenic effects. Its airborne spores are one of the potential causes of asthma in children and lung diseases in humans. The pig and chicken populations in the farms are the most affected by this fungus and its mycotoxins. Certain fungicides like mancozeb, copper oxychloride, and sulfur have inhibitory effects on the growth of this fungus and its mycotoxin producing capacities.
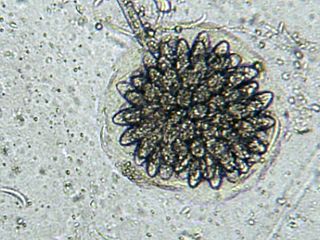
Rhopalomyces elegans is a common species of zygomycete fungus, and the type species of the genus Rhopalomyces. Widely distributed, it is found in soil, rotting plant material, and animal dung. It is a facultative parasite of nematode eggs.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.

Aspergillus versicolor is a slow-growing species of filamentous fungus commonly found in damp indoor environments and on food products. It has a characteristic musty odor associated with moldy homes and is a major producer of the hepatotoxic and carcinogenic mycotoxin sterigmatocystin. Like other Aspergillus species, A. versicolor is an eye, nose, and throat irritant.
Geomyces pannorum is a yellow-brown filamentous fungus of the phylum Ascomycota commonly found in cold soil environments including the permafrost of the Northern hemisphere. A ubiquitous soil fungus, it is the most common species of the genus Geomyces; which also includes G. vinaceus and G. asperulatus. Geomyces pannorum has been identified as an agent of disfigurement of pigments used in the 15,000-year-old paintings on the walls of the Lascaux caves of France. Strains of Geomyces have been recovered from the Alaskan Fox Permafrost Tunnel and radiocarbon dated to between 14,000 and 30,000 years old.

Cunninghamella echinulata is a fungal species in the genus Cunninghamella. It is an asexually reproducing fungus and a mesophile, preferring intermediate temperature ranges. C. echinulata is a common air contaminant, and is currently of interest to the biotechnology industry due to its ability to synthesize γ-linolenic acid as well as its capacity to bioconcentrate metals. This species is a soil saprotroph that forms rhizoids, preferring soils enriched in nitrogen, phosphorus and potassium. It has been reported occasionally an agent of mucormycosis following the inhalation of fungal spores. Czapek's agar is a suitable growth medium for the propagation of C. echinulata.

Aspergillus clavatus is a species of fungus in the genus Aspergillus with conidia dimensions 3–4.5 x 2.5–4.5 μm. It is found in soil and animal manure. The fungus was first described scientifically in 1834 by the French mycologist John Baptiste Henri Joseph Desmazières.

Cladosporium sphaerospermum is a radiotrophic fungus belonging to the genus Cladosporium and was described in 1886 by Albert Julius Otto Penzig from the decaying leaves and branches of Citrus. It is a dematiaceous (darkly-pigmented) fungus characterized by slow growth and largely asexual reproduction. Cladosporium sphaerospermum consists of a complex of poorly morphologically differentiated, "cryptic" species that share many physiological and ecological attributes. In older literature, all of these sibling species were classified as C. sphaerospermum despite their unique nature. Accordingly, there is confusion in older literature reports on the physiological and habitat regularities of C. sphaerospermum in the strict sense. This fungus is most phylogenetically similar to C. fusiforme. According to modern phylogenetic analyses, the previously synonymized species, Cladosporium langeroni, is a distinct species.
Mycorrhizal amelioration of heavy metals or pollutants is a process by which mycorrhizal fungi in a mutualistic relationship with plants can sequester toxic compounds from the environment, as a form of bioremediation.

Penicillium spinulosum is a non-branched, fast-growing fungus with a swelling at the terminal of the stipe (vesiculate) in the genus Penicillium. P. spinulosum is able to grow and reproduce in environment with low temperature and low water availability, and is known to be acidotolerant. P. spinulosum is ubiquitously distributed, and can often be isolated from soil. Each individual strain of P. spinulosum differs from others in their colony morphology, including colony texture, amount of sporulation and roughness of conidia and conidiophores.
Aspergillus wentii is an asexual, filamentous, endosymbiotic fungus belonging to the mold genus, Aspergillus. It is a common soil fungus with a cosmopolitan distribution, although it is primarily found in subtropical regions. Found on a variety of organic materials, A. wentii is known to colonize corn, cereals, moist grains, peanuts and other ground nut crops. It is also used in the manufacture of biodiesel from lipids and is known for its ability to produce enzymes used in the food industry.

Mariannaea elegans an anamorphic fungus. It is mainly found on rotting wood and soil. M. elegans is not pathogenic to humans, animals, or plants.
Botryotrichum piluliferum is a fungal species first identified in 1885 by Saccardo and Marchal. It was discovered to be the asexual state of a member of the ascomycete genus, Chaetomium. The name B. piluliferum now applies to the fungus in all its states. B. piluliferum has been found worldwide in a wide range of habitats such as animal dung and vegetation. The colonies of this fungus start off white and grow rapidly to a brown colour. The conidia are smooth and white. B. piluliferum grows optimally at a temperature of 25-30 °C and a pH of 5.5.
Microascus manginii is a filamentous fungal species in the genus Microascus. It produces both sexual (teleomorph) and asexual (anamorph) reproductive stages known as M. manginii and Scopulariopsis candida, respectively. Several synonyms appear in the literature because of taxonomic revisions and re-isolation of the species by different researchers. M. manginii is saprotrophic and commonly inhabits soil, indoor environments and decaying plant material. It is distinguishable from closely related species by its light colored and heart-shaped ascospores used for sexual reproduction. Scopulariopsis candida has been identified as the cause of some invasive infections, often in immunocompromised hosts, but is not considered a common human pathogen. There is concern about amphotericin B resistance in S. candida.
Aspergillus giganteus is a species of fungus in the genus Aspergillus that grows as a mold. It was first described in 1901 by Wehmer, and is one of six Aspergillus species from the Clavati section of the subgenus Fumigati. Its closest taxonomic relatives are Aspergillus rhizopodus and Aspergillus longivescia.
Myriodontium keratinophilum is a fungus widespread in nature, most abundantly found in keratin-rich environments such as feathers, nails and hair. Despite its ability to colonize keratinous surfaces of human body, the species has been known to be non-pathogenic in man and is phylogentically distant to other human pathogenic species, such as anthropophilic dermatophytes. However, its occasional isolation from clinical specimens along with its keratinolytic properties suggest the possibility it may contribute to disease.
Oidiodendron cereale is a species of ascomycetes fungi in the order Helotiales. This fungus is found globally in temperate climates where average summer temperatures are below 25 °C, but there have been scattered reports from tropical and subtropical environments. It is predominantly found in soil, but little is known regarding their ecological roles in nature. However, an enzymatic study from Agriculture Canada showed that O. cereale can break down a variety of plant, fungal, and animal based substrates found in soil, which may have beneficial effects for plants. On rare occasions, this fungus is found on human skin and hair. There has been one reported case of O. cereale infection in 1969, causing Neurodermitis Nuchae.