
Pro-opiomelanocortin (POMC) is a precursor polypeptide with 241 amino acid residues. POMC is synthesized in corticotrophs of the anterior pituitary from the 267-amino-acid-long polypeptide precursor pre-pro-opiomelanocortin (pre-POMC), by the removal of a 26-amino-acid-long signal peptide sequence during translation. POMC is part of the central melanocortin system.

Leptin is a protein hormone predominantly made by adipocytes. Its primary role is likely to regulate long-term energy balance.

Neuropeptide Y (NPY) is a 36 amino-acid neuropeptide that is involved in various physiological and homeostatic processes in both the central and peripheral nervous systems. It is secreted alongside other neurotransmitters such as GABA and glutamate.

The immunoglobulin heavy chain (IgH) is the large polypeptide subunit of an antibody (immunoglobulin). In human genome, the IgH gene loci are on chromosome 14.
Melanin-concentrating hormone (MCH), also known as pro-melanin stimulating hormone (PMCH), is a cyclic 19-amino acid orexigenic hypothalamic peptide originally isolated from the pituitary gland of teleost fish, where it controls skin pigmentation. In mammals it is involved in the regulation of feeding behavior, mood, sleep-wake cycle and energy balance.
GnRH2, also known as gonadotropin-releasing hormone II or LHRH-II. Its gene is located on human chromosome 20.

Urocortin is a protein that in humans is encoded by the UCN gene. Urocortin belongs to the corticotropin-releasing factor (CRF) family of proteins which includes CRF, urotensin I, sauvagine, urocortin II and urocortin III. Urocortin is involved in the mammalian stress response, and regulates aspects of appetite and stress response.

Peptide YY (PYY), also known as peptide tyrosine tyrosine, is a peptide that in humans is encoded by the PYY gene. Peptide YY is a short peptide released from cells in the ileum and colon in response to feeding. In the blood, gut, and other elements of periphery, PYY acts to reduce appetite; similarly, when injected directly into the central nervous system, PYY is also anorexigenic, i.e., it reduces appetite.

In fish anatomy and turtle anatomy, a barbel is a slender, whiskerlike sensory organ near the mouth. Fish that have barbels include the catfish, the carp, the goatfish, the hagfish, the sturgeon, the zebrafish, the black dragonfish and some species of shark such as the sawshark. Barbels house the taste buds of such fish and are used to search for food in murky water.

The Japanese rice fish, also known as the medaka, is a member of genus Oryzias (ricefish), the only genus in the subfamily Oryziinae. This small native of Japan is a denizen of rice paddies, marshes, ponds, slow-moving streams and tide pools. It is euryhaline, occurring in both brackish and freshwater. It became popular as an aquarium fish because of its hardiness and pleasant coloration: its coloration varies from creamy-white to yellowish in the wild to white, creamy-yellow, or orange in aquarium-bred individuals. Bright yellow, red or green transgenic populations, similar to GloFish, have also been developed, but are banned from sale in the EU. The medaka has been a popular pet since the 17th century in Japan. After fertilization, the female carries her eggs attached anterior to the anal fin for a period before depositing them on plants or similar things.

The type-I interferons (IFN) are cytokines which play essential roles in inflammation, immunoregulation, tumor cells recognition, and T-cell responses. In the human genome, a cluster of thirteen functional IFN genes is located at the 9p21.3 cytoband over approximately 400 kb including coding genes for IFNα, IFNω (IFNW1), IFNɛ (IFNE), IFNк (IFNK) and IFNβ (IFNB1), plus 11 IFN pseudogenes.

Leptin receptor, also known as LEP-R or OB-R, is a type I cytokine receptor, a protein that in humans is encoded by the LEPR gene. LEP-R functions as a receptor for the fat cell-specific hormone leptin. LEP-R has also been designated as CD295. Its location is the cell membrane, and it has extracellular, trans-membrane and intracellular sections.

The central melanocortin system is defined anatomically as a collection of central nervous system circuits which include:

Genetically modified fish are organisms from the taxonomic clade which includes the classes Agnatha, Chondrichthyes and Osteichthyes whose genetic material (DNA) has been altered using genetic engineering techniques. In most cases, the aim is to introduce a new trait to the fish which does not occur naturally in the species, i.e. transgenesis.
Fish are exposed to large oxygen fluctuations in their aquatic environment since the inherent properties of water can result in marked spatial and temporal differences in the concentration of oxygen. Fish respond to hypoxia with varied behavioral, physiological, and cellular responses to maintain homeostasis and organism function in an oxygen-depleted environment. The biggest challenge fish face when exposed to low oxygen conditions is maintaining metabolic energy balance, as 95% of the oxygen consumed by fish is used for ATP production releasing the chemical energy of nutrients through the mitochondrial electron transport chain. Therefore, hypoxia survival requires a coordinated response to secure more oxygen from the depleted environment and counteract the metabolic consequences of decreased ATP production at the mitochondria.

Relaxin family peptide hormones in humans are represented by seven members: three relaxin-like (RLN) and four insulin-like (INSL) peptides: RLN1, RLN2, RNL3, INSL3, INSL4, INSL5, INSL6. This subdivision into two classes is based primarily on early findings, and does not reflect the evolutionary origins or physiological differences between peptides. For example, it is known that the genes coding for RLN3 and INSL5 arose from one ancestral gene, and INSL3 shares origin with RLN2 and its multiple duplicates: RLN1, INSL4, INSL6.
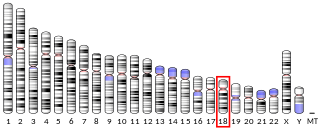
Retinal homeobox protein Rx also known as retina and anterior neural fold homeobox is a protein that in humans is encoded by the RAX gene. The RAX gene is located on chromosome 18 in humans, mice, and rats.
Stanniocalcin is a family of hormones which regulate calcium and phosphate balance in the body. The first stanniocalcin discovered was from fish and was identified as the principal calcium-reducing (hypocalcaemic) factor. It was isolated from special organs in fish called corpuscles of Stannius, hence the name stanniocalcin. Chemically, stanniocalcins are glycosylated proteins having a molecular mass of 50 kDa. They exist in molecular pairs (homodimers) and are joined together by disulfide linkage. Stanniocalcins are made up of approximately 250 amino acids.
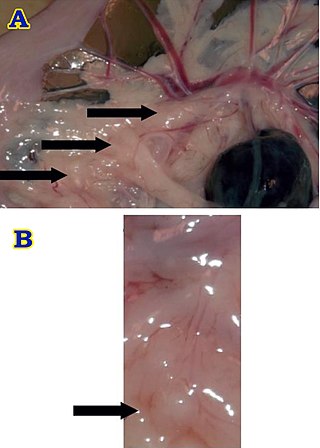
Brockmann body is an endocrine organ in some teleost fish, and is composed of a collection of islet tissues. The islet tissues are in turn composed of endocrine cells which are the principal sites of insulin synthesis. They are distributed around the spleen and the large intestine. They also secrete other hormones such as glucagon and somatostatin. Hence, Brochmann body is the centre of control of blood glucose level in these fishes. Glucagon is also produced from the intestine, but Brockmann body is the major source. Increased level of glucose stimulate the Brockmann body to release insulin, while inhibiting glucagon. Somatostatin released from Brockmann body inhibits cells to produce insulin and glucagon. In addition it inhibits release of growth hormone from the pituitary. It is named after a German physician Heinrich Brochmann who discovered it in 1848.
The food-entrainable oscillator (FEO) is a circadian clock that can be entrained by varying the time of food presentation. It was discovered when a rhythm was found in rat activity. This was called food anticipatory activity (FAA), and this is when the wheel-running activity of mice decreases after feeding, and then rapidly increases in the hours leading up to feeding. FAA appears to be present in non-mammals (pigeons/fish), but research heavily focuses on its presence in mammals. This rhythmic activity does not require the suprachiasmatic nucleus (SCN), the central circadian oscillator in mammals, implying the existence of an oscillator, the FEO, outside of the SCN, but the mechanism and location of the FEO is not yet known. There is ongoing research to investigate if the FEO is the only non-light entrainable oscillator in the body.




