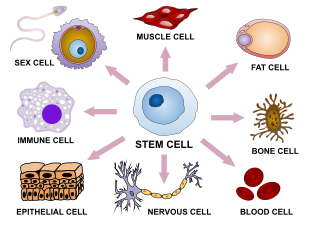
Cellular differentiation is the process in which a cell changes from one cell type to another. Usually, the cell changes to a more specialized type. Differentiation occurs numerous times during the development of a multicellular organism as it changes from a simple zygote to a complex system of tissues and cell types. Differentiation continues in adulthood as adult stem cells divide and create fully differentiated daughter cells during tissue repair and during normal cell turnover. Some differentiation occurs in response to antigen exposure. Differentiation dramatically changes a cell's size, shape, membrane potential, metabolic activity, and responsiveness to signals. These changes are largely due to highly controlled modifications in gene expression and are the study of epigenetics. With a few exceptions, cellular differentiation almost never involves a change in the DNA sequence itself. Although metabolic composition does get altered quite dramatically where stem cells are characterized by abundant metabolites with highly unsaturated structures whose levels decrease upon differentiation. Thus, different cells can have very different physical characteristics despite having the same genome.

MyoD, also known as myoblast determination protein 1, is a protein in animals that plays a major role in regulating muscle differentiation. MyoD, which was discovered in the laboratory of Harold M. Weintraub, belongs to a family of proteins known as myogenic regulatory factors (MRFs). These bHLH transcription factors act sequentially in myogenic differentiation. Vertebrate MRF family members include MyoD1, Myf5, myogenin, and MRF4 (Myf6). In non-vertebrate animals, a single MyoD protein is typically found.

Medulloblastoma is a common type of primary brain cancer in children. It originates in the part of the brain that is towards the back and the bottom, on the floor of the skull, in the cerebellum, or posterior fossa.

Bone morphogenetic protein 4 is a protein that in humans is encoded by BMP4 gene. BMP4 is found on chromosome 14q22-q23

Chemokine ligand 3 (CXCL3) is a small cytokine belonging to the CXC chemokine family that is also known as GRO3 oncogene (GRO3), GRO protein gamma (GROg) and macrophage inflammatory protein-2-beta (MIP2b). CXCL3 controls migration and adhesion of monocytes and mediates its effects on its target cell by interacting with a cell surface chemokine receptor called CXCR2. More recently, it has been shown that Cxcl3 regulates cell autonomously the migration of the precursors of cerebellar granule neurons toward the internal layers of cerebellum, during the morphogenesis of cerebellum. Moreover, if the expression of Cxcl3 is reduced in cerebellar granule neuron precursors, this highly enhances the frequency of the medulloblastoma, the tumor of cerebellum. In fact, the reduced expression of Cxcl3 forces the cerebellar granule neuron precursors to remain at the surface of the cerebellum, where they highly proliferate under the stimulus of Sonic hedgehog, becoming target of transforming insults. Remarkably, the treatment with CXCL3 completely prevents the growth of medulloblastoma lesions in a Shh-type mouse model of medulloblastoma. Thus, CXCL3 is a target for medulloblastoma therapy. Cxcl3 is directly regulated transcriptionally by BTG2

Histone deacetylase 4, also known as HDAC4, is a protein that in humans is encoded by the HDAC4 gene.

DNA-binding protein inhibitor ID-3 is a protein that in humans is encoded by the ID3 gene.

Histone deacetylase 9 is an enzyme that in humans is encoded by the HDAC9 gene.

Homeobox protein Hox-C8 is a protein that in humans is encoded by the HOXC8 gene.
Protein Tob1 is a protein that in humans is encoded by the TOB1 gene.

Homeobox protein Hox-B9 is a protein that in humans is encoded by the HOXB9 gene.
Interferon-related developmental regulator 1 is a protein that in humans is encoded by the IFRD1 gene. The gene is expressed mostly in neutrophils, skeletal and cardiac muscle, brain, pancreas. The rat and the mouse homolog genes of interferon-related developmental regulator 1 gene are also known with the name PC4 and Tis21, respectively. IFRD1 is member of a gene family that comprises a second gene, IFRD2, also known as SKmc15.

CCR4-NOT transcription complex subunit 7 is a protein that in humans is encoded by the CNOT7 gene. It is a subunit of the CCR4-Not deadenylase complex.
Protein BTG1 is a protein that in humans is encoded by the BTG1 gene.
Protein atonal homolog 1 is a protein that in humans is encoded by the ATOH1 gene.
Neurogenins are a family of bHLH transcription factors involved in specifying neuronal differentiation. It is one of many gene families related to the atonal gene in Drosophila. Other positive regulators of neuronal differentiation also expressed during early neural development include NeuroD and ASCL1.
Zinc finger protein 238 is a zinc finger containing transcription factor that in humans is encoded by the ZNF238 gene.

Protein BTG3 is a protein that in humans is encoded by the BTG3 gene.

T-box, brain, 1 is a transcription factor protein important in vertebrate embryo development. It is encoded by the TBR1 gene. This gene is also known by several other names: T-Brain 1, TBR-1, TES-56, and MGC141978. TBR1 is a member of the TBR1 subfamily of T-box family transcription factors, which share a common DNA-binding domain. Other members of the TBR1 subfamily include EOMES and TBX21. TBR1 is involved in the differentiation and migration of neurons and is required for normal brain development. TBR1 interacts with various genes and proteins in order to regulate cortical development, specifically within layer VI of the developing six-layered human cortex. Studies show that TBR1 may play a role in major neurological diseases such as Alzheimer's disease (AD), Parkinson's disease (PD) and autism spectrum disorder (ASD).

Protein BTG4 also known as BTG family member 4 is a protein that in humans is encoded by the BTG4 gene (B-cell translocation gene 4). BTG4 has anti-proliferative properties and can induce G1 cell cycle arrest.




