
The insulin receptor (IR) is a transmembrane receptor that is activated by insulin, IGF-I, IGF-II and belongs to the large class of receptor tyrosine kinase. Metabolically, the insulin receptor plays a key role in the regulation of glucose homeostasis; a functional process that under degenerate conditions may result in a range of clinical manifestations including diabetes and cancer. Insulin signalling controls access to blood glucose in body cells. When insulin falls, especially in those with high insulin sensitivity, body cells begin only to have access to lipids that do not require transport across the membrane. So, in this way, insulin is the key regulator of fat metabolism as well. Biochemically, the insulin receptor is encoded by a single gene INSR, from which alternate splicing during transcription results in either IR-A or IR-B isoforms. Downstream post-translational events of either isoform result in the formation of a proteolytically cleaved α and β subunit, which upon combination are ultimately capable of homo or hetero-dimerisation to produce the ≈320 kDa disulfide-linked transmembrane insulin receptor.

Insulin-like growth factor 1 (IGF-1), also called somatomedin C, is a hormone similar in molecular structure to insulin which plays an important role in childhood growth, and has anabolic effects in adults. In the 1950s IGF-1 was called "sulfation factor" because it stimulated sulfation of cartilage in vitro, and in the 1970s due to its effects it was termed "nonsuppressible insulin-like activity" (NSILA).

Sex hormone-binding globulin (SHBG) or sex steroid-binding globulin (SSBG) is a glycoprotein that binds to androgens and estrogens. When produced by the Sertoli cells in the seminiferous tubules of the testis, it is called androgen-binding protein (ABP).
The prolactin receptor (PRLR) is a type I cytokine receptor encoded in humans by the PRLR gene on chromosome 5p13-14. It is the receptor for prolactin (PRL). The PRLR can also bind to and be activated by growth hormone (GH) and human placental lactogen (hPL). The PRLR is expressed in the mammary glands, pituitary gland, and other tissues. It plays an important role in lobuloalveolar development of the mammary glands during pregnancy and in lactation.
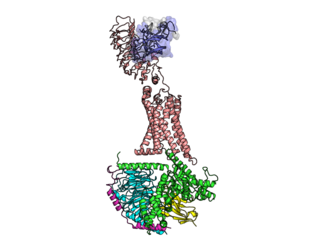
The luteinizing hormone/choriogonadotropin receptor (LHCGR), also lutropin/choriogonadotropin receptor (LCGR) or luteinizing hormone receptor (LHR), is a transmembrane receptor found predominantly in the ovary and testis, but also many extragonadal organs such as the uterus and breasts. The receptor interacts with both luteinizing hormone (LH) and chorionic gonadotropins and represents a G protein-coupled receptor (GPCR). Its activation is necessary for the hormonal functioning during reproduction.
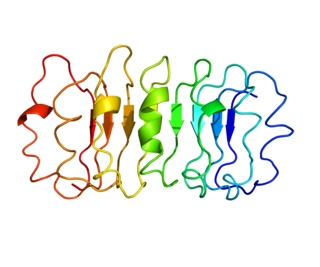
The thyrotropin receptor is a receptor that responds to thyroid-stimulating hormone and stimulates the production of thyroxine (T4) and triiodothyronine (T3). The TSH receptor is a member of the G protein-coupled receptor superfamily of integral membrane proteins and is coupled to the Gs protein.

Growth hormone receptor is a protein that in humans is encoded by the GHR gene. GHR orthologs have been identified in most mammals.

Laron syndrome (LS), also known as growth hormone insensitivity or growth hormone receptor deficiency (GHRD), is an autosomal recessive disorder characterized by a lack of insulin-like growth factor 1 production in response to growth hormone. It is usually caused by inherited growth hormone receptor (GHR) mutations.

The growth-hormone-releasing hormone receptor (GHRHR) is a G-protein-coupled receptor that binds growth hormone-releasing hormone. The GHRHR activates a Gs protein that causes a cascade of cAMP via adenylate cyclase. GHRHR is distinct from the growth hormone secretagogue receptor, where growth hormone releasing peptides act to release growth hormone.

Rev-Erb beta (Rev-Erbβ), also known as nuclear receptor subfamily 1 group D member 2 (NR1D2), is a member of the Rev-Erb protein family. Rev-Erbβ, like Rev-Erbα, belongs to the nuclear receptor superfamily of transcription factors and can modulate gene expression through binding to gene promoters. Together with Rev-Erbα, Rev-Erbβ functions as a major regulator of the circadian clock. These two proteins are partially redundant. Current research suggests that Rev-Erbβ is less important in maintaining the circadian clock than Rev-Erbα; knock-out studies of Rev-Erbα result in significant circadian disruption but the same has not been found with Rev-Erbβ. Rev-Erbβ compensation for Rev-Erbα varies across tissues, and further research is needed to elucidate the separate role of Rev-Erbβ.
Insulin-like growth factor-binding protein 3, also known as IGFBP-3, is a protein that in humans is encoded by the IGFBP3 gene. IGFBP-3 is one of six IGF binding proteins that have highly conserved structures and bind the insulin-like growth factors IGF-1 and IGF-2 with high affinity. IGFBP-7, sometimes included in this family, shares neither the conserved structural features nor the high IGF affinity. Instead, IGFBP-7 binds IGF1R, which blocks IGF-1 and IGF-2 binding, resulting in apoptosis.

Parathyroid hormone/parathyroid hormone-related peptide receptor, also known as parathyroid hormone 1 receptor (PTH1R), is a protein that in humans is encoded by the PTH1R gene. PTH1R functions as a receptor for parathyroid hormone (PTH) and for parathyroid hormone-related protein (PTHrP), also called parathyroid hormone-like hormone (PTHLH).

Somatostatin receptor type 2 is a protein that in humans is encoded by the SSTR2 gene.

Corticotropin-releasing hormone receptor 1 (CRHR1) is a protein, also known as CRF1, with the latter (CRF1) now being the IUPHAR-recommended name. In humans, CRF1 is encoded by the CRHR1 gene at region 17q21.31, beside micrototubule-associated protein tau MAPT.

Thyroid hormone receptor beta (TR-beta) also known as nuclear receptor subfamily 1, group A, member 2 (NR1A2), is a nuclear receptor protein that in humans is encoded by the THRB gene.
Corticotropin-releasing hormone receptor 2 (CRHR2) is a protein, also known by the IUPHAR-recommended name CRF2, that is encoded by the CRHR2 gene and occurs on the surfaces of some mammalian cells. CRF2 receptors are type 2 G protein-coupled receptors for corticotropin-releasing hormone (CRH) that are resident in the plasma membranes of hormone-sensitive cells. CRH, a peptide of 41 amino acids synthesized in the hypothalamus, is the principal neuroregulator of the hypothalamic-pituitary-adrenal axis, signaling via guanine nucleotide-binding proteins (G proteins) and downstream effectors such as adenylate cyclase. The CRF2 receptor is a multi-pass membrane protein with a transmembrane domain composed of seven helices arranged in a V-shape. CRF2 receptors are activated by two structurally similar peptides, urocortin II, and urocortin III, as well as CRH.

Growth hormone 2 (GH2), also known more commonly as placental growth hormone (PGH) or growth hormone variant (GH-V), is a protein that in humans is encoded by the GH2 gene. It is produced by and secreted from the placenta during pregnancy, and becomes the predominant form of growth hormone (GH) in the body during this time. Its cogener is growth hormone 1 (GH1), or pituitary growth hormone.

Thyroid stimulating hormone, beta also known as TSHB is a protein which in humans is encoded by the TSHB gene.

Kowarski syndrome describes cases of growth failure, despite the presence of normal or slightly high blood growth hormone by radioimmunoassay (RIA-GH) and low serum IGF1, and who exhibit a significant increase in growth rate following recombinant GH therapy.

Examorelin (INN) (developmental code names EP-23905, MF-6003), also known as hexarelin, is a potent, synthetic, peptidic, orally-active, centrally-penetrant, and highly selective agonist of the ghrelin/growth hormone secretagogue receptor (GHSR) and a growth hormone secretagogue which was developed by Mediolanum Farmaceutici. It is a hexapeptide with the amino acid sequence His-D-2-methyl-Trp-Ala-Trp-D-Phe-Lys-NH2 which was derived from GHRP-6. These GH-releasing peptides have no sequence similarity to ghrelin, but mimic ghrelin by acting as agonists at the ghrelin receptor.


