Paleobiota
Color key
| Notes Uncertain or tentative taxa are in small text; |
Synapsids
An egg-like object was discovered by Llewellyn Price in the vicinity of Rattlesnake Canyon, and subsequently described by Romer and Price in 1939. It had a shell-like texture which was not easily comparable to nodules from the Permian of Texas, and the authors considered it to have potentially been laid by a "pelycosaur". If this identification is correct, it is the oldest amniotic egg in the fossil record. [9] [1] Later investigations concluded that a distinct calcareous shell layer was not present, though the high concentration of phosphorus in the object suggests that it may still be an egg, albeit one with a softer outer membrane. [10] [3]
| Synapsids of the Nocona Formation | ||||
|---|---|---|---|---|
| Genus | Species | Localities | Notes | Images |
| Ctenorhachis [11] | C. jacksoni [11] | Lake Kickapoo [11] | A large sphenacodontid with a rather low neural spine sail. |  |
| Dimetrodon | D. booneorum | Briar Creek [1] [12] | A medium-sized sphenacodontid of uncertain validity. | |
| D. limbatus [1] | Briar Creek?, [1] [12] [13] Coprolite, Loftin, Rattlesnake Canyon 2 [3] | A large sphenacodontid with a tall neural spine sail. |  | |
| D. natalis | Geraldine, [2] [8] Briar Creek, [1] [12] [13] Rattlesnake Canyon 2 [3] | A small sphenacodontid with a tall neural spine sail. Represented by three partial skeletons. Sometimes regarded as a juvenile of D. limbatus, [2] [3] but confirmed to be a unique species with small adults, according to histological analyses. [12] |  | |
| Edaphosaurus | E. boanerges [1] | Geraldine, [1] [2] [14] [8] Briar Creek, [1] [13] Coprolite, Loftin, Rattlesnake Canyon 2? [3] | A medium-sized herbivorous edaphosaurid. [1] [14] One of the most common and characteristic fossils of the Geraldine bonebed, with multiple articulated skeletons displayed in museums around the world. [2] |  |
| Lupeosaurus | L. kayi | Briar Creek, [1] Coprolite? [3] | A large edaphosaurid known from rare fragments. |  |
| Ophiacodon | O. retroversus | Rattlesnake Canyon [1] [15] | An ophiacodontid. |  |
| O. uniformis | (vicinity of) Geraldine, [2] [8] Briar Creek [1] [15] | An ophiacodontid. |  | |
| Secodontosaurus | S. obtusidens | Briar Creek [1] [12] | A medium-sized sphenacodontid. |  |
Reptiles
| Reptiles of the Nocona Formation | ||||
|---|---|---|---|---|
| Genus | Species | Localities | Notes | Images |
| Araeoscelis | A. casei [16] | Godwin Creek [16] | An araeoscelidian initially named as a new genus, Ophiodeirus. [16] |  |
| Bolosaurus | B. striatus | Geraldine, [2] [8] Briar Creek, Godwin Creek [17] | A bolosaurid parareptile. |  |
| Captorhinidae indet. | Loftin [3] | Rare captorhinid vertebrae. [18] [3] | ||
Amphibians
| Amphibians of the Nocona Formation | ||||
|---|---|---|---|---|
| Genus | Species | Localities | Notes | Images |
| Archeria | A. crassidisca | Geraldine, [19] [2] [14] [8] Briar Creek, [20] Coprolite, Loftin, Rattlesnake Canyon 2 [3] | A large archeriid embolomere. Multiple articulated skeletons have been found in the Geraldine bonebed, representing the most complete and well-described fossils of this species. [19] [2] [14] |  |
| Cardiocephalus | C. sp. | Geraldine [8] | Rare microsaur teeth. | |
| Diadectes | D. sideropelicus | Geraldine, [2] Briar Creek, [20] Coprolite, Loftin, Rattlesnake Canyon 2 [3] | A large diadectid diadectomorph. |  |
| Diplocaulus | D. sp. | Loftin [3] | A diplocaulid nectridean. | |
| Eryops | E. megacephalus | Geraldine, [2] [8] Briar Creek, [20] Coprolite, Loftin, Rattlesnake Canyon 2 [3] | An eryopid, a type of large semiaquatic temnospondyl. Articulated skeletons and other remains are common in the Geraldine bonebed. [2] | 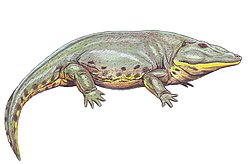 |
| Rubeostratilia [21] | R. texensis [21] | "east of Henrietta" [21] | An amphibamiform, a type of small terrestrial temnospondyl. |  |
| Scapanops [22] | S. neglecta [22] | Halsell Hill [22] | A eucacopine dissorophid, a type of small terrestrial temnospondyl. Previously consider a specimen of Conjunctio . [22] | |
| Trimerorhachis | T. insignis | (vicinity of) Geraldine, [2] [8] Godwin Creek, [23] Rattlesnake Canyon, [23] Loftin, Rattlesnake Canyon 2 [3] [23] | A trimerorhachid dvinosaur, a type of small aquatic temnospondyl. |  |
| Zatrachys | Z. serratus | (vicinity of) Geraldine, [2] Rattlesnake Canyon 2 | A zatracheid temnospondyl, a type of medium-sized terrestrial temnospondyl. |  |
Fish
Indeterminate palaeoniscoids are known from the Geraldine, [8] Coprolite, Loftin, and Rattlesnake Canyon 2 bonebeds. [3] Iniopterygian tooth whorls have been reported from the Rattlesnake Canyon area. [8]
| Fish of the Nocona Formation | ||||
|---|---|---|---|---|
| Genus | Species | Localities | Notes | Images |
| Acanthodes | A. sp. | Geraldine [8] | Acanthodian fin spines and scales |  |
| Barbclabornia | B. luedersensis | Rattlesnake Canyon [24] | Rare xenacanth shark teeth | |
| Ectosteorhachis | E. nitidus | Geraldine, [2] Coprolite, Loftin [3] | A megalichthyid tetrapodomorph |  |
| Helodus | H. sp. | Geraldine [8] | Rare holocephalan teeth | |
| Hybodus | H. sp. | Coprolite [3] | A hybodont shark | |
| Janassa? | J.? sp. | Geraldine [8] | A single petalodont tooth | |
| Orthacanthus | O. platypternus | Geraldine [8] | Xenacanth shark teeth |  |
| O. texensis | Geraldine, [2] [8] Briar Creek, [8] Coprolite, [25] Loftin, Rattlesnake Canyon 2 [3] | Very common xenacanth shark teeth and coprolites | ||
| Platysomus? | P.? sp. | Geraldine [8] | A single palaeoniscoid tooth | |
| Progyrolepis | P. tricessimalaris [26] | Rattlesnake Canyon [26] | A palaeoniscid known from a partial skeleton. | |
| Sagenodus | S. periprion | (vicinity of) Geraldine, [2] [8] Coprolite, Rattlesnake Canyon 2 [3] | Lungfish teeth | |
| Spermatodus | S. pustulosus | Rattlesnake Canyon 2 [3] | A coelacanth | |
| Xenacanthus | X. sp. | Geraldine [8] | Xenacanth shark spine fragments |  |
Plants
Plant fossils are known from several bonebeds of the Nocona Formation, though they are subordinate to vertebrate fossils at most sites. [27] [2] Insect damage has been recorded on leaves from the Coprolite Bonebed. It is uncommon (only a third as frequent as in modern plants), even when compared to only slightly younger sites such as the Taint locality in the Waggoner Ranch Formation. Despite the rarity of insect damage, the Coprolite Bonebed shows the oldest occurrence of skeletonization (removal of all but the veins), as well as galls, which are rarely found in Permian plant fossils. [28]
| Plants of the Nocona Formation | ||||
|---|---|---|---|---|
| Genus | Species | Localities | Notes | Images |
| Annularia | A. cf. stellata | Geraldine, [2] Coprolite [28] | Foliage of a large calamitacean sphenophyte (horsetail) which formed thickets in shallow water and shorelines. | |
| Autunia | A. cf. conferta | Geraldine, [2] Coprolite [28] | Foliage of a peltasperm "seed fern", previously considered a species of Callipteris. [2] Abundant in the Geraldine and Coprolite bonebeds. [2] [28] | |
| Calamites | C. undulatus | Geraldine, [2] Coprolite [3] [28] | Common stem impressions of a large calamitacean sphenophyte (horsetail) which formed thickets in shallow water and shorelines. | |
| Callipteridium | C. cf. pteridium | Geraldine [2] | Foliage of a medullosalean "seed fern". | |
| C. virginianum | Geraldine [2] | Foliage of a medullosalean "seed fern". | ||
| Cordaites | C. principalis | Geraldine, [2] Coprolite [28] | Foliage of a cordaitalean gymnosperm, a small tree found in both swamps and uplands. Common in the Geraldine and Coprolite bonebeds. [2] [28] | |
| Dadoxylon | D. sp. | Geraldine, [2] Rattlesnake Canyon 2 [3] | Conifer wood and charcoal. | |
| Odontopteris | O. genuina | Geraldine [2] | Foliage of a medullosalean "seed fern". | |
| O. cf. lingulata | Coprolite [3] | Foliage of a medullosalean "seed fern". | ||
| O. cf. osmundaeformis | Geraldine [2] | Foliage of a medullosalean "seed fern". | ||
| Pecopteris | P. arborescens | Geraldine [2] | Foliage of a marattialean tree fern, among the most common trees in Permian swamp environments. | |
| P. candolleana | Geraldine [2] | |||
| P. hemitelioides | Geraldine, [2] Coprolite [3] [28] | |||
| P. unita | Geraldine [2] | |||
| Psaronius | P. sp. | Geraldine, [2] Loftin [3] | Stems and roots of a marattialean tree fern, likely the same plant which produced Pecopteris leaves. [3] | |
| Russellites [29] | R. taeniata | Geraldine, [2] Coprolite [28] | Foliage of a cycadophyte, previously referred to Tingia. Common in the Coprolite Bonebed. [28] | |
| Samaropsis | S. spp. | Geraldine [2] | Two species of gymnosperm seeds, possibly from the same plants as Walchia and Cordaites. [2] | |
| Sigillaria | S. brardii | Geraldine [2] | Bark and stems of a lycophyte. | |
| Sphenophyllum | S. oblongifolium | Geraldine [2] | Foliage of a fern. | |
| Sphenopteris | S. cf. macilenta | Geraldine [2] | Foliage of a fern or "seed fern". | |
| Walchia | W. piniformis | Geraldine, [2] Coprolite [3] [28] | Foliage of an early conifer, a small tree accustomed to dry uplands. Associated stems and charcoal show similarity to the form genus Schizodendron. [3] Very common in the Geraldine and Coprolite bonebeds. [2] [28] | |