| Archer City Formation | |
|---|---|
| Stratigraphic range: Early Permian, Asselian–early Sakmarian | |
| Type | Formation |
| Unit of | Bowie Group |
| Underlies | Nocona Formation |
| Overlies | Markley Formation |
| Lithology | |
| Primary | mudstone |
| Other | sandstone, siltstone |
| Location | |
| Region | |
| Country | |
| Type section | |
| Named for | Archer City, Texas |
| Named by | Hentz & Brown, 1987 |
 | |
The Archer City Formation is a geological formation in north-central Texas, preserving fossils from the Asselian and early Sakmarian stages of the Permian period. It is the earliest component of the Texas red beds, introducing a tropical ecosystem which will persist in the area through the rest of the Early Permian. The Archer City Formation is preceded by the cool Carboniferous swamp sediments of the Markley Formation, and succeeded by the equally fossiliferous red beds of the Nocona Formation. The Archer City Formation was not named as a unique geological unit until the late 1980s. Older studies generally labelled its outcrops as the Moran or Putnam formations, which are age-equivalent marine units to the southwest.
Contents
- History and stratigraphy
- Age
- Paleoenvironment
- Geology
- Sandstone beds
- Paleosols
- Paleobiota
- Synapsids
- Reptiles
- Temnospondyls
- Other amphibians
- Fish
- Plants
- See also
- References
The sediments of the Archer City Formation reconstruct a coastal floodplain crossed by small meandering rivers and isolated ponds. [1] [2] The climate was hot and monsoonal, preserving a variety of paleosols and plant assemblages ranging from dry upland forests to moist riverside galleries. "Seed ferns" ( Sphenopteris , Autunia , Odontopteris , etc.) and early conifers ( Walchia , etc.) occupied drier soils, while marattialean ferns ( Pecopteris , Asterotheca , etc.) and horsetails ( Sphenophyllum , Annularia , Calamites , etc.) were prominent in more humid areas.
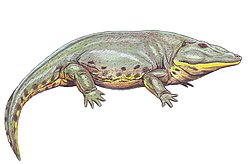
The Archer City Formation preserves a diverse fauna of fossil vertebrates in pond bonebeds across Archer and Clay counties. Many fossils are rare fragments of species which are better-preserved in younger parts of the red beds. Nevertheless, complete skeletons and important type specimens are also known from the formation. Some notable synapsids include Apsisaurus witteri , Edaphosaurus boanerges , Ophiacodon uniformis , and Dimetrodon milleri . Early reptiles such as Protorothyris archeri and Romeria spp. make up a small component of the terrestrial fauna. Amphibians are common in every niche, with examples such as Phonerpeton pricei, Neldasaurus wrightei , Edops craigi, Eryops megacephalus , and Pantylus cordatus , just to name a few. The rivers were roamed by Orthacanthus texensis (a xenacanth "shark") and Ectosteorhachis nitidus (a megalichthyid fish).