This enzyme belongs to the family of isomerases, specifically those intramolecular oxidoreductases interconverting aldoses and ketoses. The systematic name of this enzyme class is N-(5-phospho-beta-D-ribosyl)anthranilate aldose-ketose-isomerase. Other names in common use include:
PRA isomerase,
PRAI,
IGPS:PRAI (indole-3-glycerol-phosphate,
synthetase/N-5'-phosphoribosylanthranilate isomerase complex), and
Phosphoribosylanthranilate isomerase is one of the many enzymes within the biosynthesis pathway of tryptophan (an essential amino acid). The upstream* pathway substrates and intermediates are shown below (Fig. 2).
As seen in Fig. 3, N-(5'-phosphoribosyl)-anthranilate via this enzyme is converted into 1-(o-carboxyphenylamino)-1-deoxribulose 5-phosphate. As the name phosphoribosylanthranilate isomerase suggests, it functions as an isomerase, rearranging the parts of the molecule without adding or removing molecules or atoms.
The reaction seen in Fig. 3, is an intramolecular redox (reduction-oxidation) reaction.[5] Its first step involves a proton transfer. This product intermediate, an enolamine, is fluorescent, which is useful for kinetic studies
within this pathway.[5] However, this product is unstable, and quickly isomerases into an α-amino keto.
Fig. 2: Upstream* Pathway of Tryptophan Synthesis
Fig 3: Enzyme Isomerase Reaction
Fig. 4: Downstream* Pathway of Tryptophan Synthesis
Note: Upstream/Downstream are relative to the compounds/molecules directly involved in phosphoribosylanthranilate isomerase reaction
Kinetics
Michaelis–Menten kinetics data, is given in the table below for PRAI and indole-glycerol-phosphate synthase (IGPS, EC 4.1.1.48).[6]
Table 1: Kinetic Data
Enzyme
Temperature (°C)
Km
(μM)
kcat
(1/sec)
tPRAI
25
0.280
3.7
45
0.390
13.5
60
0.730
38.5
80
1.030
116.8
tIGPS
25
0.006
0.11
45
0.014
0.75
60
0.053
3.24
80
0.123
15.4
Structure
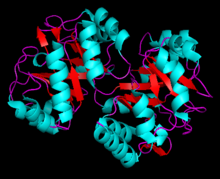
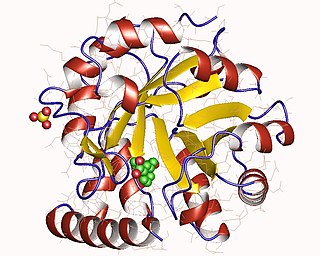
Fig 6: Structure of N-(5'-phosphoribosyl) anthranilate isomerase from Pyrococcus furiosus
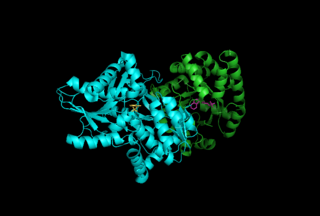
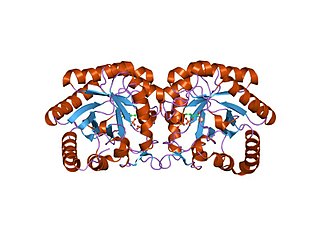
Depending on the microorganism PRAI's structure can vary between a mono-functional enzyme (monomeric and labile) or a stable bi-functional dimeric enzyme. Within Saccharomyces cerevisiae, Bacillus subtilis, Pseudomonas putida, and Acinetobacter calcoaceticus the enzyme is monmeric.[7] In contrast, in hyperthermophileThermotoga maritima,Escherichia coli (Fig. 5), Salmonella typhimurium, and Aerobacter aerogenes, and Serratia marcescens, it is a bi-functional enzyme with indoleglycerol phosphate synthase as the paired enzyme.[8]
The crystal structure has been characterized for a variety of the above listed microorganisms. The known 2.0 A structure of PRAI from Pyrococcus furiosus shows that tPRAI has a TIM-barrel fold (Fig. 6). PRAI derived from Thermococcus kodakaraensis also expresses a similar TIM-barrel fold structure.[7] The subunits of tPRAI associate via the N-terminal faces of their central beta-barrels. Two long, symmetry-related loops that protrude reciprocally into cavities of the other subunit provide for multiple hydrophobic interactions. Moreover, the side chains of the N-terminal methionines and the C-terminal leucines of both subunits are immobilized in a hydrophobic cluster, and the number of salt bridges is increased in tPRAI. These features appear to be mainly responsible for the high thermostability of tPRAI.[9]
Fig 5: Three dimensional structure of the bi-functional PRAI: IGPS enzyme from E. Coli Fig 7: IGPS (purple), shared (orange), and PRAI (turquoise) reaction domains
The bi-functional version of this enzyme isolated from E. Coli (Fig. 5) performs two steps within the Tryptophan pathway. Referencing Fig. 7, the N-terminal catalyzes the IGPS reaction (residues ~1–289 purple), and the C-terminal domain performs the PRAI reaction (residues ~158–452 turquoise). Although these domains overlap (orange), the active sites are not overlapping, and studies have shown that mono-functional enzymes composing of these two domains are still able to produce a functional tryptophan bio-synthetic pathway.[10]
The βα loops are responsible for the activity of this enzyme, and the αβ loops are involved in the protein's stability.[8]
More details on the discovery of this enzyme's structure can be found in Willmann's paper.[11]
Specifically, for phosphoribosyl anthranilate isomerase, TkTrpF, from Thermococcus kodakaraensis. The active site for the Amadori rearrangement, was determined to be Cys8 (acting as the general base) and Asp135 (as the general acid).[12]
Inhibitors
An enzyme inhibitor[13] is molecule that binds to an enzyme that therefore decreases the activity of the protein. The following molecules have been shown to inhibit PRAI activity:
Reduced 1-(2-carboxyphenylamino )-1-deoxy-D-ribulose 5-phosphate [5, 6,8); Indoleglycerol phosphate (8); Indolepropanol phosphate (8); MnCI2 CoCI2 [16); CuS04 (16); More (chemically synthesized N-(5-phospho-betaD-ribosyl)anthranilate contains inhibitors, but not if it is generated by anthranilate phosphoribosyltransferase)
A list of genes encoding for PRAI can also be found on KEGG Enzyme database.[15]
Related Research Articles
Tryptophan synthase or tryptophan synthetase is an enzyme that catalyzes the final two steps in the biosynthesis of tryptophan. It is commonly found in Eubacteria, Archaebacteria, Protista, Fungi, and Plantae. However, it is absent from Animalia. It is typically found as an α2β2 tetramer. The α subunits catalyze the reversible formation of indole and glyceraldehyde-3-phosphate (G3P) from indole-3-glycerol phosphate (IGP). The β subunits catalyze the irreversible condensation of indole and serine to form tryptophan in a pyridoxal phosphate (PLP) dependent reaction. Each α active site is connected to a β active site by a 25 Ångstrom long hydrophobic channel contained within the enzyme. This facilitates the diffusion of indole formed at α active sites directly to β active sites in a process known as substrate channeling. The active sites of tryptophan synthase are allosterically coupled.
Antimycins are produced as secondary metabolites by Streptomyces bacteria, a soil bacteria. These specialized metabolites likely function to kill neighboring organisms in order to provide the streptomyces bacteria with a competitive edge.
Shikimic acid, more commonly known as its anionic form shikimate, is a cyclohexene, a cyclitol and a cyclohexanecarboxylic acid. It is an important biochemical metabolite in plants and microorganisms. Its name comes from the Japanese flower shikimi, from which it was first isolated in 1885 by Johan Fredrik Eykman. The elucidation of its structure was made nearly 50 years later.
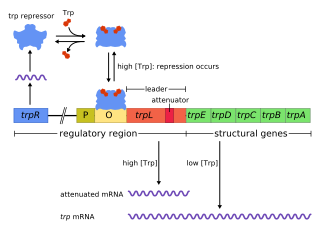
The trp operon is a group of genes that are transcribed together, encoding the enzymes that produce the amino acid tryptophan in bacteria. The trp operon was first characterized in Escherichia coli, and it has since been discovered in many other bacteria. The operon is regulated so that, when tryptophan is present in the environment, the genes for tryptophan synthesis are repressed.
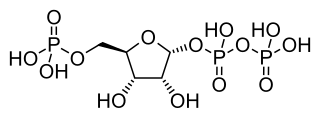
Phosphoribosyl pyrophosphate (PRPP) is a pentose phosphate. It is a biochemical intermediate in the formation of purine nucleotides via inosine-5-monophosphate, as well as in pyrimidine nucleotide formation. Hence it is a building block for DNA and RNA. The vitamins thiamine and cobalamin, and the amino acid tryptophan also contain fragments derived from PRPP. It is formed from ribose 5-phosphate (R5P) by the enzyme ribose-phosphate diphosphokinase:
Amino acid biosynthesis is the set of biochemical processes by which the amino acids are produced. The substrates for these processes are various compounds in the organism's diet or growth media. Not all organisms are able to synthesize all amino acids. For example, humans can synthesize 11 of the 20 standard amino acids. These 11 are called the non-essential amino acids).
Phosphoribosylformylglycinamidine cyclo-ligase is the fifth enzyme in the de novo synthesis of purine nucleotides. It catalyzes the reaction to form 5-aminoimidazole ribotide (AIR) from formylglycinamidine-ribonucleotide FGAM. This reaction closes the ring and produces a 5-membered imidazole ring of the purine nucleus (AIR):
The enzyme anthranilate synthase catalyzes the chemical reaction
The enzyme indole-3-glycerol-phosphate synthase (IGPS) (EC 4.1.1.48) catalyzes the chemical reaction
In enzymology, an aminodeoxychorismate synthase is an enzyme that catalyzes the chemical reaction
In molecular biology, the protein domain SAICAR synthase is an enzyme which catalyses a reaction to create SAICAR. In enzymology, this enzyme is also known as phosphoribosylaminoimidazolesuccinocarboxamide synthase. It is an enzyme that catalyzes the chemical reaction
The enzyme 3-dehydroquinate synthase catalyzes the chemical reaction
The enzyme methylglyoxal synthase catalyzes the chemical reaction
In enzymology, an anthranilate phosphoribosyltransferase is an enzyme that catalyzes the chemical reaction
5′-Phosphoribosyl-5-aminoimidazole is a biochemical intermediate in the formation of purine nucleotides via inosine-5-monophosphate, and hence is a building block for DNA and RNA. The vitamins thiamine and cobalamin also contain fragments derived from AIR. It is an intermediate in the adenine pathway and is synthesized from 5′-phosphoribosylformylglycinamidine by AIR synthetase.
3-Deoxy-D-arabinoheptulosonate 7-phosphate (DAHP) synthase is the first enzyme in a series of metabolic reactions known as the shikimate pathway, which is responsible for the biosynthesis of the amino acids phenylalanine, tyrosine, and tryptophan. Since it is the first enzyme in the shikimate pathway, it controls the amount of carbon entering the pathway. Enzyme inhibition is the primary method of regulating the amount of carbon entering the pathway. Forms of this enzyme differ between organisms, but can be considered DAHP synthase based upon the reaction that is catalyzed by this enzyme.
5-enolpyruvylshikimate-3-phosphate (EPSP) synthase is an enzyme produced by plants and microorganisms. EPSPS catalyzes the chemical reaction:
3-Deoxy-D-arabino-heptulosonic acid 7-phosphate (DAHP) is a 7-carbon ulosonic acid. This compound is found in the shikimic acid biosynthesis pathway and is an intermediate in the production of aromatic amino acids.
Tryptophan synthase (indole-salvaging) (EC 4.2.1.122, tryptophan synthase beta2) is an enzyme with systematic name L-serine hydro-lyase (adding indole, L-tryptophan-forming). This enzyme catalyses the following chemical reaction
Li Jiayang is a Chinese agronomist and geneticist. He is Vice Minister of Agriculture in China and President of the Chinese Academy of Agricultural Sciences (CAAS). He is also Professor and Principal investigator at the Institute of Genetics and Development at the Chinese Academy of Sciences (CAS).
References
↑ Creighton TE, Yanofsky C (1970). "Chorismate to tryptophan (Escherichia coli)—anthranilate synthetase, PR transferase, PRA isomerase, InGP synthetase, tryptophan synthetase". Metabolism of Amino Acids and Amines Part A. Methods in Enzymology. Vol.17A. pp.365–380. doi:10.1016/0076-6879(71)17215-1. ISBN9780121818746.
1 2 Hommel U, Eberhard M, Kirschner K (April 1995). "Phosphoribosyl anthranilate isomerase catalyzes a reversible amadori reaction". Biochemistry. 34 (16): 5429–39. doi:10.1021/bi00016a014. PMID7727401.
↑ Sterner R, Merz A, Thoma R, Kirschner K (2001). "Phosphoribosylanthranilate isomerase and indoleglycerol-phosphate synthase: Tryptophan biosynthetic enzymes from Thermotoga maritima". Hyperthermophilic enzymes Part B. Methods in Enzymology. Vol.331. pp.270–80. doi:10.1016/S0076-6879(01)31064-9. ISBN9780121822323. PMID11265469.
↑ Hennig M, Sterner R, Kirschner K, Jansonius JN (May 1997). "Crystal structure at 2.0 A resolution of phosphoribosyl anthranilate isomerase from the hyperthermophile Thermotoga maritima: possible determinants of protein stability". Biochemistry. 36 (20): 6009–16. doi:10.1021/bi962718q. PMID9166771.
↑ Eberhard M, Tsai-Pflugfelder M, Bolewska K, Hommel U, Kirschner K (April 1995). "Indoleglycerol phosphate synthase-phosphoribosyl anthranilate isomerase: comparison of the bifunctional enzyme from Escherichia coli with engineered monofunctional domains". Biochemistry. 34 (16): 5419–28. doi:10.1021/bi00016a013. PMID7727400.
↑ PDB: 1PII; Wilmanns M, Priestle JP, Niermann T, Jansonius JN (January 1992). "Three-dimensional structure of the bifunctional enzyme phosphoribosylanthranilate isomerase: indoleglycerolphosphate synthase from Escherichia coli refined at 2.0 A resolution". Journal of Molecular Biology. 223 (2): 477–507. doi:10.1016/0022-2836(92)90665-7. PMID1738159.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.
 Fig. 2: Upstream* Pathway of Tryptophan Synthesis
Fig. 2: Upstream* Pathway of Tryptophan Synthesis Fig 3: Enzyme Isomerase Reaction
Fig 3: Enzyme Isomerase Reaction Fig. 4: Downstream* Pathway of Tryptophan Synthesis
Fig. 4: Downstream* Pathway of Tryptophan Synthesis