
Papilio polyxenes, the (eastern) black swallowtail, American swallowtail or parsnip swallowtail, is a butterfly found throughout much of North America. An extremely similar-appearing species, Papilio joanae, occurs in the Ozark Mountains region, but it appears to be closely related to Papilio machaon, rather than P. polyxenes. The species is named after the figure in Greek mythology, Polyxena, who was the youngest daughter of King Priam of Troy. Its caterpillar is called the parsley worm because the caterpillar feeds on parsley.

The small heath is a butterfly species belonging to the family Nymphalidae, classified within the subfamily Satyrinae. It is the smallest butterfly in this subfamily. The small heath is diurnal and flies with a noticeable fluttering flight pattern near the ground. It rests with closed wings when not in flight. It is widespread in colonies throughout the grasslands of Eurasia and north-western Africa, preferring drier habitats than other Coenonympha, such as salt marshes, alpine meadows, wetlands, and grasslands near water. However, habitat loss caused by human activities has led to a decline in populations in some locations.

Gonepteryx rhamni, commonly named the common brimstone, is a butterfly of the family Pieridae. It lives throughout the Palearctic zone and is commonly found across Europe, Asia, and North Africa. Across much of its range, it is the only species of its genus, and is therefore simply known locally as the brimstone. Its wing span size is 60–74 mm (2.4–2.9 in). It should not be confused with the brimstone moth Opisthograptis luteolata.

A spermatophore or sperm ampulla is a capsule or mass containing spermatozoa created by males of various animal species, especially salamanders and arthropods, and transferred in entirety to the female's ovipore during reproduction. Spermatophores may additionally contain nourishment for the female, in which case it is called a nuptial gift, as in the instance of bush crickets. In the case of the toxic moth Utetheisa ornatrix, the spermatophore includes sperm, nutrients, and pyrrolizidine alkaloids which prevent predation because it is poisonous to most organisms. However, in some species such as the Edith's checkerspot butterfly, the "gift" provides little nutrient value. The weight of the spermatophore transferred at mating has little effect on female reproductive output.

Diplacus aurantiacus, the sticky monkey-flower or orange bush monkey-flower, is a flowering plant that grows in a subshrub form, native to southwestern North America from southwestern Oregon south through most of California. It is a member of the lopseed family, Phrymaceae. It was formerly known as Mimulus aurantiacus.

Polygonia c-album, the comma, is a food generalist (polyphagous) butterfly species belonging to the family Nymphalidae. The angular notches on the edges of the forewings are characteristic of the genus Polygonia, which is why species in the genus are commonly referred to as anglewing butterflies. Comma butterflies can be identified by their prominent orange and dark brown/black dorsal wings.

Heliconius charithonia, the zebra longwing or zebra heliconian, is a species of butterfly belonging to the subfamily Heliconiinae of the family Nymphalidae. It was first described by Carl Linnaeus in his 1767 12th edition of Systema Naturae. The boldly striped black and white wing pattern is aposematic, warning off predators. And it’s the state butterfly of Florida.

The marsh fritillary is a butterfly of the family Nymphalidae. Commonly distributed in the Palearctic region, the marsh fritillary's common name derives from one of its several habitats, marshland. The prolonged larval stage lasts for approximately seven to eight months and includes a period of hibernation over the winter. The larvae are dependent on the host food plant Succisa pratensis not only for feeding but also for hibernation, because silken webs are formed on the host plant as the gregarious larvae enter hibernation. Females lay eggs in batches on the host plant and are, like other batch-layers, selective about the location of oviposition because offspring survivorship levels for batch-layers are more tied to location selection than they are for single-egg layers.

The queen butterfly is a North and South American butterfly in the family Nymphalidae with a wingspan of 80–85 mm. It is orange or brown with black wing borders and small white forewing spots on its dorsal wing surface, and reddish ventral wing surface fairly similar to the dorsal surface. The ventral hindwings have black veins and small white spots in a black border. The male has a black androconial scent patch on its dorsal hindwings. It can be found in meadows, fields, marshes, deserts, and at the edges of forests.

Papilio demodocus, the citrus swallowtail or Christmas butterfly, is a swallowtail butterfly which commonly occurs over the entirety of sub-Saharan Africa, including Madagascar, besides the southern Arabian Peninsula. The caterpillars feed on various native plants of especially the family Rutaceae, but have also taken to the leaves of cultivated citrus trees.

Scrophularia californica is a flowering plant in the figwort family which is known by the common names California figwort and California bee plant.

Epargyreus clarus, the silver-spotted skipper, is a butterfly of the family Hesperiidae. It is claimed to be the most recognized skipper in North America. E. clarus occurs in fields, gardens, and at forest edges and ranges from southern Canada throughout most of the United States to northern Mexico, but is absent in the Great Basin and western Texas.

Jalmenus evagoras, the imperial hairstreak, imperial blue, or common imperial blue, is a small, metallic blue butterfly of the family Lycaenidae. It is commonly found in eastern coastal regions of Australia. This species is notable for its unique mutualism with ants of the genus Iridomyrmex. The ants provide protection for juveniles and cues for adult mating behavior. They are compensated with food secreted from J. evagoras larvae. The ants greatly enhance the survival and reproductive success of the butterflies. J. evagoras lives and feeds on Acacia plants, so butterfly populations are localized to areas with preferred species of both host plants and ants.

Utetheisa ornatrix, also called the ornate bella moth, ornate moth, bella moth or rattlebox moth, is a moth of the subfamily Arctiinae. It is aposematically colored ranging from pink, red, orange and yellow to white coloration with black markings arranged in varying patterns on its wings. It has a wingspan of 33–46 mm. Moths reside in temperate midwestern and eastern North America as well as throughout Mexico and other parts of Central America. Unlike most moths, the bella moth is diurnal. Formerly, the bella moth or beautiful utetheisa of temperate eastern North America was separated as Utetheisa bella. Now it is united with the bella moth in Utetheisa ornatrix.
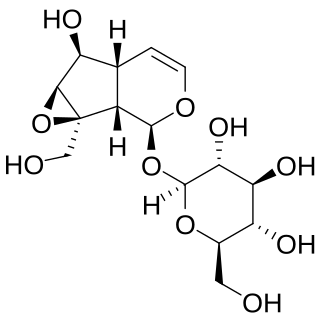
Catalpol is an iridoid glucoside. This natural product falls in the class of iridoid glycosides, which are simply monoterpenes with a glucose molecule attached.

The Quino checkerspot is a butterfly native to southern California and northwestern Baja California. It is a subspecies of the common Edith's checkerspot and the second such subspecies to be listed under the federal Endangered Species Act.

Edith's checkerspot is a species of butterfly in the family Nymphalidae. It is a resident species of western North America and among the subspecies, entomologists have long been intrigued by their many phenotypic variations in coloration, wing length, and overall body size. Most populations are monophagous and rely on plants including Plantago erecta and Orthocarpus densiflorus as their host species in developing from eggs through to larvae, pupae, and mature butterflies. Males exhibit polygyny whereas females rarely mate more than once. Males devote most of their attention to mate acquisition, and such mate locating strategies such as hilltopping behavior have developed. Climate change and habitat destruction have impacted certain subspecies. Three subspecies in particular, Euphydryas editha quino, Euphydryas editha bayensis and Euphydryas editha taylori, are currently under protection via the Endangered Species Act.

Bicyclus anynana is a small brown butterfly in the family Nymphalidae, the most globally diverse family of butterflies. It is primarily found in eastern Africa from southern Sudan to Eswatini. It is found mostly in woodland areas and flies close to the ground. Male wingspans are 35–40 mm and female wingspans are 45–49 mm.

Euphydryas editha taylori, the Whulge checkerspot or Taylor's checkerspot, is a butterfly native to an area of the northwestern United States and Vancouver Island.

The Bay checkerspot is a butterfly endemic to the San Francisco Bay region of the U.S. state of California. It is a federally threatened species, as a subspecies of Euphydryas editha.



