Related Research Articles

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.

The cerebral cortex, also known as the cerebral mantle, is the outer layer of neural tissue of the cerebrum of the brain in humans and other mammals. It is the largest site of neural integration in the central nervous system, and plays a key role in attention, perception, awareness, thought, memory, language, and consciousness. The cerebral cortex is the part of the brain responsible for cognition.

In neuroanatomy, the trigeminal nerve (lit. triplet nerve), also known as the fifth cranial nerve, cranial nerve V, or simply CN V, is a cranial nerve responsible for sensation in the face and motor functions such as biting and chewing; it is the most complex of the cranial nerves. Its name (trigeminal, from Latin tri- 'three' and -geminus 'twin') derives from each of the two nerves (one on each side of the pons) having three major branches: the ophthalmic nerve (V1), the maxillary nerve (V2), and the mandibular nerve (V3). The ophthalmic and maxillary nerves are purely sensory, whereas the mandibular nerve supplies motor as well as sensory (or "cutaneous") functions. Adding to the complexity of this nerve is that autonomic nerve fibers as well as special sensory fibers (taste) are contained within it.

The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.
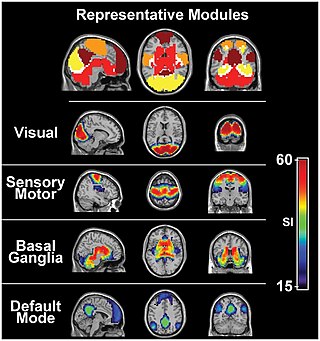
The sensory nervous system is a part of the nervous system responsible for processing sensory information. A sensory system consists of sensory neurons, neural pathways, and parts of the brain involved in sensory perception and interoception. Commonly recognized sensory systems are those for vision, hearing, touch, taste, smell, balance and visceral sensation. Sense organs are transducers that convert data from the outer physical world to the realm of the mind where people interpret the information, creating their perception of the world around them.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.

A Brodmann area is a region of the cerebral cortex, in the human or other primate brain, defined by its cytoarchitecture, or histological structure and organization of cells. The concept was first introduced by the German anatomist Korbinian Brodmann in the early 20th century. Brodmann mapped the human brain based on the varied cellular structure across the cortex and identified 52 distinct regions, which he numbered 1 to 52. These regions, or Brodmann areas, correspond with diverse functions including sensation, motor control, and cognition.
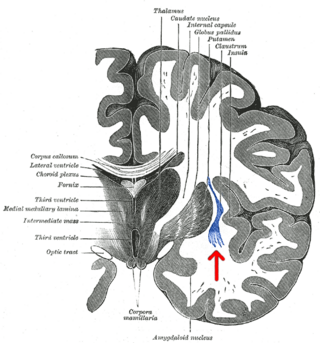
The claustrum is a thin sheet of neurons and supporting glial cells in the brain, that connects to the cerebral cortex and subcortical regions including the amygdala, hippocampus and thalamus. It is located between the insular cortex laterally and the putamen medially, encased by the extreme and external capsules respectively. Blood to the claustrum is supplied by the middle cerebral artery. It is considered to be the most densely connected structure in the brain, and thus hypothesized to allow for the integration of various cortical inputs such as vision, sound and touch, into one experience. Other hypotheses suggest that the claustrum plays a role in salience processing, to direct attention towards the most behaviorally relevant stimuli amongst the background noise. The claustrum is difficult to study given the limited number of individuals with claustral lesions and the poor resolution of neuroimaging.

Sensory neurons, also known as afferent neurons, are neurons in the nervous system, that convert a specific type of stimulus, via their receptors, into action potentials or graded receptor potentials. This process is called sensory transduction. The cell bodies of the sensory neurons are located in the dorsal root ganglia of the spinal cord.
The receptive field, or sensory space, is a delimited medium where some physiological stimuli can evoke a sensory neuronal response in specific organisms.

In neuroanatomy, the superior colliculus is a structure lying on the roof of the mammalian midbrain. In non-mammalian vertebrates, the homologous structure is known as the optic tectum or optic lobe. The adjective form tectal is commonly used for both structures.

In neuroanatomy, thalamocortical radiations, also known as thalamocortical fibers, are the efferent fibers that project from the thalamus to distinct areas of the cerebral cortex. They form fiber bundles that emerge from the lateral surface of the thalamus.

Retinotopy is the mapping of visual input from the retina to neurons, particularly those neurons within the visual stream. For clarity, 'retinotopy' can be replaced with 'retinal mapping', and 'retinotopic' with 'retinally mapped'.
In medicine and anatomy, the special senses are the senses that have specialized organs devoted to them:

A bipolar neuron, or bipolar cell, is a type of neuron characterized by having both an axon and a dendrite extending from the soma in opposite directions. These neurons are predominantly found in the retina and olfactory system. The embryological period encompassing weeks seven through eight marks the commencement of bipolar neuron development.
The isothalamus is a division used by some researchers in describing the thalamus.
A sensory map is an area of the brain which responds to sensory stimulation, and are spatially organized according to some feature of the sensory stimulation. In some cases the sensory map is simply a topographic representation of a sensory surface such as the skin, cochlea, or retina. In other cases it represents other stimulus properties resulting from neuronal computation and is generally ordered in a manner that reflects the periphery. An example is the somatosensory map which is a projection of the skin's surface in the brain that arranges the processing of tactile sensation. This type of somatotopic map is the most common, possibly because it allows for physically neighboring areas of the brain to react to physically similar stimuli in the periphery or because it allows for greater motor control.
Feature detection is a process by which the nervous system sorts or filters complex natural stimuli in order to extract behaviorally relevant cues that have a high probability of being associated with important objects or organisms in their environment, as opposed to irrelevant background or noise.

In the human brain, the nucleus basalis, also known as the nucleus basalis of Meynert or nucleus basalis magnocellularis, is a group of neurons located mainly in the substantia innominata of the basal forebrain. Most neurons of the nucleus basalis are rich in the neurotransmitter acetylcholine, and they have widespread projections to the neocortex and other brain structures.

Michel Imbert is a neuropsychologist teacher-researcher in cognitive neurosciences, professor emeritus at Pierre-et-Marie-Curie University and honorary director of studies at the École des hautes études en sciences sociales (EHESS).
References
- ↑ Hinds, O.; et al. (2009). "Locating the functional and anatomical boundaries of human primary visual cortex". NeuroImage. 46 (4): 915–922. doi:10.1016/j.neuroimage.2009.03.036. PMC 2712139 . PMID 19328238.
- ↑ Udin, S.; Fawcett, J. (1988). "Formation of Topographic Maps". Annu. Rev. Neurosci. 11: 289–327. doi:10.1146/annurev.ne.11.030188.001445. PMID 3284443.
- ↑ Rosa, M.G.P. (2002). "Visual maps in the adult primate cerebral cortex: some implications for brain development and evolution". Braz J Med Biol Res. 35 (12): 1485–1498. doi: 10.1590/s0100-879x2002001200008 . PMID 12436190.
- ↑ Talavage, T. M.; et al. (2000). "Frequency-dependent responses exhibited by multiple regions in human auditory cortex". Hearing Research. 150 (1–2): 225–244. doi:10.1016/s0378-5955(00)00203-3. PMID 11077206. S2CID 8678128.
- ↑ Dimitrijevic, M. R., et al. (2005). Motor Control in the Human Spinal Cord. Artificial Organs. 29(3): 216-219.
- ↑ Zullo L, Sumbre G, Agnisola C, Flash T, Hochner B. (2009). Nonsomatotopic organization of the higher motor centers in octopus. Curr Biol. 19(19):1632-6.
- 1 2 Auffarth, B (2013). "Understanding smell -- the olfactory stimulus problem". Neuroscience & Biobehavioral Reviews. 37 (8): 1667–1679. doi:10.1016/j.neubiorev.2013.06.009. PMID 23806440. S2CID 207090474.
- ↑ Imai, T.; et al. (2010). "Topographic Mapping—The Olfactory System". Cold Spring Harb Perspect Biol. 2 (8): 8. doi:10.1101/cshperspect.a001776. PMC 2908763 . PMID 20554703.
- ↑ Vosshall, L.; et al. (2000). "An Olfactory Sensory Map in the Fly Brain". Cell. 102 (2): 147–159. doi: 10.1016/s0092-8674(00)00021-0 . PMID 10943836. S2CID 17573876.
- ↑ Wang, F; Nemes, A; Meldelson, M; Axel, R (1998). "Odorant Receptors Govern the Formation of a Precise Topographic Map". Cell. 93 (1): 47–60. doi: 10.1016/s0092-8674(00)81145-9 . PMID 9546391. S2CID 18458277.
- ↑ Kaas, J. H., Qi, H. X., & Iyengar, S. (2006). Cortical network for representing the teeth and tongue in primates. Anatomical Record Part a-Discoveries in Molecular Cellular and Evolutionary Biology, 288A(2): 182-190.
- ↑ Zaidi, F. N.; Whitehead, M. C. (2006). "Discrete Innervation of Murine Taste Buds by Peripheral Taste Neurons". J. Neurosci. 26 (32): 8243–8253. doi: 10.1523/jneurosci.5142-05.2006 . PMC 6673808 . PMID 16899719.
- ↑ Kaas, J. H. (2008). "The evolution of the complex sensory and motor systems of the human brain". Brain Research Bulletin. 75 (2–4): 384–390. doi:10.1016/j.brainresbull.2007.10.009. PMC 2349093 . PMID 18331903.
- ↑ Kaas, J. H. (1997). "Topographic Maps are Fundamental to Sensory Processing". Brain Research Bulletin. 44 (2): 107–112. doi:10.1016/s0361-9230(97)00094-4. PMID 9292198. S2CID 14488006.
- ↑ Schneider, KA; Richter, MC; Kastner, S (2004). "Retinotopic Organization and Functional Subdivision of the Human Lateral Geniculate Nucleus: A High-Resolution Functional Magnetic Resonance Imaging Study". J. Neurosci. 24 (41): 8975–8985. doi: 10.1523/jneurosci.2413-04.2004 . PMC 6730047 . PMID 15483116.