
Equidae is the taxonomic family of horses and related animals, including the extant horses, asses, and zebras, and many other species known only from fossils. The family evolved around 50 million years ago from a small, multi-toed ungulate into larger, single-toed animals. All extant species are in the genus Equus, which originated in North America. Equidae belongs to the order Perissodactyla, which includes the extant tapirs and rhinoceros, and several extinct families. It is more specifically grouped within the superfamily Equoidea, the only other family being the extinct Palaeotheriidae.

The molars or molar teeth are large, flat teeth at the back of the mouth. They are more developed in mammals. They are used primarily to grind food during chewing. The name molar derives from Latin, molaris dens, meaning "millstone tooth", from mola, millstone and dens, tooth. Molars show a great deal of diversity in size and shape across the mammal groups. The third molar of humans is sometimes vestigial.

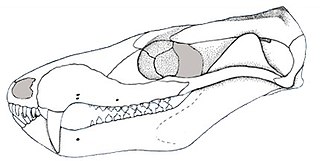
Palaeotherium is an extinct genus of equoid that lived in Europe and possibly the Middle East from the Middle Eocene to the Early Oligocene. It is the type genus of the Palaeotheriidae, a group exclusive to the Palaeogene that was closest in relation to the Equidae, which contains horses plus their closest relatives and ancestors. Fossils of Palaeotherium were first described in 1782 by the French naturalist Robert de Lamanon and then closely studied by another French naturalist, Georges Cuvier, after 1798. Cuvier erected the genus in 1804 and recognized multiple species based on overall fossil sizes and forms. As one of the first fossil genera to be recognized with official taxonomic authority, it is recognized as an important milestone within the field of palaeontology. The research by early naturalists on Palaeotherium contributed to the developing ideas of evolution, extinction, and succession and demonstrating the morphological diversity of different species within one genus.

Numidotherium is an extinct genus of early proboscideans, discovered in 1984, that lived during the middle Eocene of North Africa some 46 million years ago. It was about 90–100 cm (35–39 in) tall at the shoulder and weighed about 250–300 kg (550–660 lb).

Palaeotheriidae is an extinct family of herbivorous perissodactyl mammals that inhabited Europe, with less abundant remains also known from Asia, from the mid-Eocene to the early Oligocene. They are classified in Equoidea, along with the living family Equidae.

Anoplotherium is the type genus of the extinct Palaeogene artiodactyl family Anoplotheriidae, which was endemic to Western Europe. It lived from the late Eocene to the earliest Oligocene. It was the fifth fossil mammal genus to be described with official taxonomic authority, with a history extending back to 1804 when its fossils from Montmartre in Paris, France were first described by the French naturalist Georges Cuvier. Discoveries of incomplete skeletons of A. commune in 1807 led Cuvier to thoroughly describe unusual features for which there are no modern analogues. His drawn skeletal and muscle reconstructions of A. commune in 1812 were amongst the first instances of anatomical reconstructions based on fossil evidence. Cuvier's contributions to palaeontology based on his works on the genus were revolutionary for the field, not only proving the developing ideas of extinction and ecological succession but also paving the way for subfields such as palaeoneurology. Today, there are four known species.
Brasilodon is an extinct genus of small, mammal-like cynodonts that lived in what is now Brazil during the Norian age of the Late Triassic epoch, about 225.42 million years ago. While no complete skeletons have been found, the length of Brasilodon has been estimated at 12 centimetres (4.7 in). Its dentition shows that it was most likely an insectivore. The genus is monotypic, containing only the species B. quadrangularis. Brasilodon belongs to the family Brasilodontidae, whose members were some of the closest relatives of mammals, the only cynodonts alive today. Two other brasilodontid genera, Brasilitherium and Minicynodon, are now considered to be junior synonyms of Brasilodon.

Hipposideros besaoka is an extinct bat from Madagascar in the genus Hipposideros. It is known from numerous jaws and teeth, which were collected in a cave at Anjohibe in 1996 and described as a new species in 2007. The site where H. besaoka was found is at most 10,000 years old; other parts of the cave have yielded H. commersoni, a living species of Hipposideros from Madagascar, and some material that is distinct from both species. H. besaoka was larger than H. commersoni, making it the largest insectivorous bat of Madagascar, and had broader molars and a more robust lower jaw. As usual in Hipposideros, the second upper premolar is small and displaced from the toothrow, and the second lower premolar is large.

Triaenops goodmani is an extinct bat from Madagascar in the genus Triaenops. It is known from three lower jaws collected in a cave at Anjohibe in 1996, and described as a new species in 2007. The material is at most 10,000 years old. A bat humerus from the same site could not be identified as either T. goodmani or the living T. menamena. T. goodmani is identifiable as a member of Triaenops or the related genus Paratriaenops by a number of features of the teeth, such as the single-cusped, canine-like fourth premolar and the presence of a gap between the entoconid and hypoconulid cusps on the first two molars. T. goodmani is larger than the living species of Triaenops and Paratriaenops on Madagascar, and on the first molar the protoconid cusp is only slightly higher than the hypoconid, not much higher as in the other species.

Brontotheriidae is a family of extinct mammals belonging to the order Perissodactyla, the order that includes horses, rhinoceroses, and tapirs. Superficially, they looked rather like rhinos with some developing bony nose horns, and were some of the earliest mammals to have evolved large body sizes of several tonnes. They lived around 56–34 million years ago, until the very close of the Eocene. Brontotheres had a Holarctic distribution, with the exception of Western Europe: they occupied North America, Asia, and Eastern Europe. They were the first fossilized mammals to be discovered west of the Mississippi, and were first discovered in South Dakota.

Xiphodon is the type genus of the extinct Palaeogene artiodactyl family Xiphodontidae. It, like other xiphodonts, was endemic to Western Europe and lived from the middle Eocene up to the earliest Oligocene. Fossils from Montmartre in Paris, France that belonged to X. gracilis were first described by the French naturalist Georges Cuvier in 1804. Although he assigned the species to Anoplotherium, he recognized that it differed from A. commune by its dentition and limb bones, later moving it to its own subgenus in 1822. Xiphodon was promoted to genus rank by other naturalists in later decades. It is today defined by the type species X. gracilis and two other species, X. castrensis and X. intermedium.

Homogalax is an extinct genus of tapir-like odd-toed ungulate. It was described on the basis of several fossil finds from the northwest of the United States, whereby the majority of the remains come from the state of Wyoming. The finds date to the Lower Eocene between 56 and 48 million years ago. In general, Homogalax was very small, only reaching the weight of today's peccaries, with a maximum of 15 kg. Phylogenetic analysis suggests the genus to be a basal member of the clade that includes today's rhinoceros and tapirs. In contrast to these, Homogalax was adapted to fast locomotion.

Plagiolophus is an extinct genus of equoids belonging to the family Palaeotheriidae. It lived in Europe from the middle Oligocene to the early Oligocene. The type species P. minor was initially described by the French naturalist Georges Cuvier in 1804 based on postcranial material including a now-lost skeleton originally from the Paris Basin. It was classified to Palaeotherium the same year but was reclassified to the subgenus Plagiolophus, named by Auguste Pomel in 1847. Plagiolophus was promoted to genus rank by subsequent palaeontologists and today includes as many as seventeen species. As proposed by the French palaeontologist Jean A. Remy in 2004, it is defined by three subgenera: Plagiolophus, Paloplotherium, and Fraasiolophus.

Megaconus is an extinct genus of allotherian mammal from the Middle Jurassic Tiaojishan Formation of Inner Mongolia, China. The type and only species, Megaconus mammaliaformis was first described in the journal Nature in 2013. Megaconus is thought to have been a herbivore that lived on the ground, having a similar posture to modern-day armadillos and rock hyraxes. Megaconus was in its initial description found to be member of a group called Haramiyida. A phylogenetic analysis published along its description suggested that haramiyidans originated before the appearance of true mammals, but in contrast, the later description of the haramiyidan Arboroharamiya in the same issue of Nature indicated that haramyidans were true mammals. If haramiyidans are not mammals, Megaconus would be one of the most basal ("primitive") mammaliaforms to possess fur, and an indicator that fur evolved in the ancestors of mammals and not the mammals themselves. However, later studies cast doubt on the euharamiyidan intrepretation, instead finding it to be a basal allotherian mammal.

Dichodon is an extinct genus of Palaeogene artiodactyls belonging to the family Xiphodontidae. It was endemic to Western Europe and lived from the middle Eocene up to the earliest Oligocene. The genus was first erected by the British naturalist Richard Owen in 1848 based on dental remains from the fossil beds in Hordle, England. He noticed similar dentitions to contemporary artiodactyls like those of the Anoplotheriidae and Dichobunidae and references the name of the genus Dichobune. Eventually, it was found to be more closely related to Xiphodon and now includes 11 species, although one of them may be synonymous.

Prosotherium is an extinct genus of late Oligocene hegetotheriid notoungulate. It has been found in two Argentinian fossil formations, ie, the Sarmiento Formation in Chubut Province, and the Agua de la Piedra Formation in Mendoza Province.

Periptychus is an extinct genus of mammal belonging to the family Periptychidae. It lived from the Early to Late Paleocene and its fossil remains have been found in North America.

Diplobune is an extinct genus of Palaeogene artiodactyls belonging to the family Anoplotheriidae. It was endemic to Europe and lived from the late Eocene to the early Oligocene. The genus was first erected as a subgenus of Dichobune by Ludwig Rütimeyer in 1862 based on his hypothesis of the taxon being a transitional form between "Anoplotherium" secundaria, previously erected by Georges Cuvier in 1822, and Dichobune. He based the genus etymology off of the two-pointed pillarlike shapes of the lower molars, which had since been a diagnosis of it. However, in 1870, Diplobune was elevated to genus rank by Oscar Fraas, who recognized that Diplobune was a distinct genus related to Anoplotherium and not Dichobune. After several revisions of the anoplotheriids, there are currently four known species of which D. minor is the type species.

Dacrytherium is an extinct genus of Palaeogene artiodactyls belonging to the family Anoplotheriidae. It occurred from the Middle to Late Eocene of Western Europe and is the type genus of the subfamily Dacrytheriinae, the older of the two anoplotheriid subfamilies. Dacrytherium was first erected in 1876 by the French palaeontologist Henri Filhol, who recognised in his studies that it had dentition similar to the anoplotheriids Anoplotherium and Diplobune but differed from them by a deep preorbital fossa and a lacrimal fossa, the latter of which is where the genus name derives from. D. ovinum, originally classified in Dichobune, is the type species of Dacrytherium. Henri Filhol named D. elegans in 1884, and Hans Georg Stehlin named the species D. priscum and D. saturnini in 1910.

The research history of Palaeotherium is complicated given its extensive fossil record and lengthy taxonomic history, with the earliest record of its fossils dating back to 1782 when the French physicist Robert de Lamanon described the skull of what the naturalist Georges Cuvier described as belonging to P. medium in 1804. Cuvier initially recognized its affinities to tapirs and rhinoceroses and classified fossil material to three different species based on size. From 1805 to 1824, he established additional species based on the morphologies of postcranial remains and drew a reconstructed skeleton of P. magnum in 1824. The fossil mammal genus was the fourth to have been recognized with undisputed taxonomic authority. Palaeotherium had since been a subject of significant attention by many other palaeontologists, and it was gradually revised to be recognized as taxonomically distinct from its other perissodactyl relatives.















