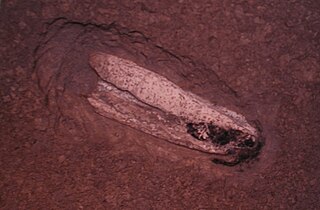
Belemnotheutis is an extinct coleoidcephalopod genus from the middle and upper Jurassic, related to but morphologically distinct from belemnites. Belemnotheutisfossils are some of the best preserved among coleoids. Remains of soft tissue are well-documented in some specimens, even down to microscopic muscle tissue. In 2008, a group of paleontologists even recovered viable ink from ink sacs found in several specimens.
This genus was the subject of a dispute between several eminent 19th century British paleontologists, notably between Richard Owen and Gideon Mantell. Some authors incorrectly spell the genus Belemnoteuthis following the usual spelling teuthis (τευθίς) for 'squid'.
Description
The genus Belemnotheutis is characterized by an internal shell consisting of a conical phragmocone covered apically by a thin rostrum, or guard, homologous to the bullet-shaped rostrum of true belemnites, a short forward projecting proostracum, and ten hook bearing arms of equal length.
Belemnotheutis fossils are sometimes found in remarkable states of preservation, some specimens retaining permineralized soft tissue.[1] The mantle, fins, head, arms, and hooks are well-documented from remains preserved in Lagerstätten. One specimen recovered from Christian Malford, Wiltshire and currently displayed in the Paleontology Department of the Natural History Museum in London is fossilized clasping a fish.[2]
Parts of the internal skeleton of Belemnotheutis (rostrum is cut away in this illustration).
The apical portion of the Belemnotheutis internal skeleton is called the rostrum (plural: rostra) or the guard. The rostrum of Belemnotheutis differs significantly from that of true belemnites. Unlike the bullet-shaped dense guards of belemnites, the rostrum of Belemnotheutis is only present as a very thin sheath. It was also composed of aragonite rather than the heavy calcite of belemnites.[5] In large specimens the rostrum can reach a maximum of only 1mm (0.039in) in thickness near the tip. The outer surface was covered by a thin organic layer in the live animal.[6] In true belemnites, the large dense rostra acted as a counterbalance, keeping the animal horizontally oriented when swimming.[7] It was long assumed that Belemnotheutis were confined to shallow waters, unable to venture into deeper waters due to the absence of the heavy rostra. The discovery of cameral deposits in the phragmocones of Belemnotheutis in 1952 made it clear that they were capable of controlling buoyancy.[7]
The phragmocone of Belemnotheutis is short and blunt, measuring around 35mm (1.4in) to 86mm (3.4in) in length.[4][9] The outer wall of the phragmocone is called the conotheca, distinct from the rostrum. It begins approximately 50mm (2.0in) from the tip of the phragmocone and consists of a nacreous outer layer and an inner lamellar layer.[10] The outer layer gradually thins from 0.6mm (0.024in) in thickness to only about 0.1mm (0.0039in) thick at about 65mm (2.6in) further down the shell until it eventually disappears around the opening of the phragmocone (the peristome). Sometimes there is a hollow gap between the rostrum and the lamellar layer of the conotheca, indicating either organic content that have since disappeared or disintegration of the lamellar layer itself.[6] The phragmocone of Belemnotheutis had about 50 chambers that were originally aragonitic, though they are usually replaced by calcium phosphate during the process of fossilization.[9][11]
At the very tip of the phragmocone beneath the rostrum is an embryonic shell known as the protoconch. In Belemnotheutis, like in other belemnotheutids, the protoconch is roughly cup-shaped and sealed.[7][12] This was thought to be another method of distinguishing it from other belemnites which usually have ball-shaped protoconchs.[12] However, it is probably a taphonomic artefact, with the protoconch being spherical like other belemnites.[13]
Reconstruction of Belemnotheutis in life
The long, weakly tapering structure in the dorsal anterior part of the internal skeleton is called the proostracum. It is striated longitudinally and often shows minute holes left by boring organisms usually less than 1μm in diameter.[6] The length of the proostracum is one to two times the length of the phragmocone. The proostracum was a thin, delicate structure substantially narrower than the phragmocone.[14] Its original composition is unknown and it is always found separated from the phragmocone and the rostrum and often crushed.[9][14] Whether the proostracum was connected to or derived from the phragmocone is still a subject of debate among paleontologists.[6] Its general morphology, however, resembles that of true belemnites rather than those from other 'unusual' belemnoid coeloids with short rostra like Permoteuthis and Phragmoteuthis.[7]
The head is not well preserved in known specimens. It comprised approximately 20% of the body length (excluding the arms). Brain cartilage is observed in some specimens, as well as a pair of aragonitic statoliths which helped the animal determine horizontal orientation when swimming.[9][14]Belemnotheutis, like most of the other belemnoids, had ten arms of equal length lined with two rows of inward-pointing hooks each.[15][16] Each of the hooks were composed of several sections. The curved pointed tip is called the uncinus and was probably the only part of the hooks exposed. The rest of the hook (the shaft and the base) were embedded in a sheath of soft tissue below the orbicular scar, a small groove where the tissue attachment terminated. They are also believed to have been stalked and mobile, helping the animal manipulate its prey.[15] Traces of functional suckers have been found and these constitute a single row on each arm, alternating in between the pairs of hooks. The size of the suckers decreases distally along the arms, with the largest (around 2mm (0.079in) in diameter) being closest to the head.[9][17] The length of the arms varies with the size of the individual but may have reached 100mm (3.9in) in larger specimens.[9]
Belemnotheutis had a cylindriconical muscularmantle covered by an outer and inner skin (tunic).[9][14] Traces of a criss-cross pattern composed of collagen fibers have been observed in the surface of some well-preserved fossils.[18] The cross section of the exceptionally preserved body wall of a specimen from the Oxford Clayformations also reveals alternating bands of concentrically and radially oriented body fibers. They imply that Belemnotheutis were powerful swimmers and agile predators, similar to modern shallow-water squid species.[19] The animal reached 10 to 30cm (3.9 to 11.8in) in length, including its arms. The body diameter was around 12 to 14% of the mantle length.[9][20] At the center of the dorsal surface of the rostrum is a narrow V-shaped groove running about 3/5ths the length of the phragmocone from the apex, with two rounded ridges at its left and right sides. These grooves are one of the most distinctive features of the Belemnotheutidae and are theorized to have served as attachments to terminal oval or oar-shaped fins like in some modern squids.[21] The siphuncle is marginal and located ventrally.[9] Directly in front of the phragmocone was an ink sac that could reach 25mm (0.98in) long in large specimens.[22] Intestinal casts (cololites) as well as the orientations and positions of fossilized remains reveal that the animal preyed on fish and other coleoids in life. Their great abundance in certain formations indicate that Belemnotheutis were highly gregarious animals, congregating in large monospecific or polyspecific shoals.[14]
Belemnotheutis are coleoids belonging to the familyBelemnotheutidae. Belemnotheutis and other belemnotheutids are considered by some paleontologists to be distinct from true belemnites (suborderBelemnitina). Most authorities like Jeletzky (1966),[7] Bandel and Kulicki (1988), and Peter Doyle (1990)[13] classify it under Belemnitida in the suborder Belemnotheutina (the classification used by this article). Others like Donovan (1977) and Engeser and Reitner (1981) classify it as a distinct order, Belemnotheutida, based on the aragonitic constitution of the rostra,[9] the shape of the proostraca, protoconchs, and the arm crowns, among other morphological factors.[12]
Belemnotheutis has been continually spelled as Belemnoteuthis by authors who believed that Pearce had made an honest mistake in naming the specimens. In 1999, D.T. Donovan and M.D. Crane succeeded in convincing the International Commission on Zoological Nomenclature that the spelling was intentional, citing historical usage of the spelling Greekθευτίς (theutis) as a valid variant of the usual τευθίς (teuthis, 'squid'). Subsequently, the accepted spelling is now formally Belemnotheutis.[9][25]
Species
The following is a list of species described under the genus Belemnotheutis.[26][27]
Fossilized ink sacs were first discovered in belemnites in 1826 by Mary Anning a famous British fossil collector and paleontologist, who along with her brother Joseph and a friend and fellow fossil collector Elizabeth Philpot succeeded in recovering the ink, used to illustrate ichthyosaur and pterosaur fossils.[30][31] The ink recovered from such fossils were also used to draw fossil ichthyosaurs by Henry De la Beche, a friend and supporter of Mary Anning.[32]
In 2008, an excavation team led by the British Geological Survey in Christian Malford recovered fossilized ink sacs from several remarkably preserved remains of Belemnotheutis antiquus in the Oxford Clay that had been previously identified during the 1840s.[19] The specimens were fossilized rapidly in apatite (calcium phosphate) through a process paleontologist Phil Wilby called "The Medusa Effect".[33] By mixing it with ammonia solution, the team was able to return the ink to its liquid form. Bringing to mind the 19th century practices of the aforementioned early paleontologists, they used the ~150 million year old ink to draw a replica of the original illustration of Belemnotheutis as drawn by Joseph Pearce.[34] Dr. Wilby called the drawing "the ultimate self-portrait".[34]
History and controversy
Left and Center: erroneous reconstructions by Richard Owen.
Right: a restoration by Joseph Pearce, the original discoverer of Belemnotheutis. It is also the illustration copied in 2008 using ~150 million year old ink recovered from the fossilized ink sac of Belemnotheutis antiquus.
In 1843, Richard Owen acquired specimens of Belemnotheutis from the same locality from another paleontologist, Samuel Pratt. He formally published a paper in 1844 (On the Belemnites, Philosophical Transactions of the Royal Society),[35] naming the specimens Belemnites oweniiPratt, after himself and crediting Pratt with the discovery while failing to mention Pearce.[36] He believed that the specimens were of the genus Belemnites whose typically lengthy rostra simply got separated. He sent a copy of the paper to Pearce in the same year, proving that he was actually aware of Pearce's earlier description but had deliberately omitted any mention of him. Pearce responded by stating that examination by another paleontologist James Bowerbank, supported his belief that fossils did not possess the bullet-shaped guards typical of Belemnites but instead had rostra in the form of very thin sheaths. Bowerbanks confirmed this assertion but supported Owen's assignment of Belemnites, saying that the presence of very short rostra did not justify the classification of Belemnotheutis as a separate genus from Belemnites.[9]
Owen received a Royal Medal from the Royal Society in 1846 for the 1844 paper,[36] further inducing Pearce to protest what he viewed as erroneous descriptions of the specimens. In 1847, the London Geological Journal published a paper by Pearce of his objections to Owen's paper. At the same time the editor of the paper and another paleontologist, Edward Charlesworth, published an editorial criticizing Owen for deliberately failing to credit Pearce with the discovery of Belemnotheutis, as well as his apparent disregard to the opinions of less well-known paleontologists like Pearce. This was also the first time that Pearce described the specific epithetantiquus to the fossils. Pearce died later in the same year in May 1847 taking no further part in what was to become a controversy. Shortly after his death, the same paper published the support of William Cunnington, a fossil collector, for this description as opposed to Owen's conclusions.[9]
In 1848, Gideon Mantell read a description of Belemnotheutis specimens recovered by his son Reginald Neville Mantell to the Royal Society.[8] His descriptions supported that of Pearce's views and held that the differences between belemnites and Belemnotheutis were enough to justify it being a separate genus. He also described the characteristic groove on the apical dorsal surface of the Belemnotheutis for the first time (structures which Owen had attributed as artifacts of crushing). He had expected Owen, who was present during the session, to support this amendment. Instead, Owen ridiculed Mantell, further aggravating the famous feud between the two.[9][37]
Mantell continued to assert his position until his death in 1852,[35] gaining supporters in other eminent paleontologists like Edward Forbes and Charles Lyell against Owen with regards to the true morphology of Belemnotheutis.[37] By then the hostility between Owen and Mantell had escalated, Owen going so far as to oppose the awarding of the Royal Medal to Mantell[36] for his work in 1849. Mantell did eventually receive the Royal Medal for his work on Iguanodon to which Owen had attempted to claim another authority much in the same way that he had named Belemnotheutis after himself.[9]
In 1860, three years after Mantell's death, Owen eventually published an amendment to his earlier descriptions. He acknowledged that Belemnotheutis indeed had very thin rostra and was distinct from the genus Belemnites. He did so only after other prominent authorities described the very similar Acanthoteuthis and were considering Belemnotheutis as its synonym.[38][39][40] However, he never recanted his earlier criticism of both Pearce and Mantell.[9]
Related Research Articles
Coleoidea or Dibranchiata is one of the two subclasses of cephalopods containing all the various taxa popularly thought of as "soft-bodied" or "shell-less". Unlike its extant sister group Nautiloidea, whose members have a rigid outer shell for protection, the coleoids have at most an internal shell called cuttlebone or gladius that is used for buoyancy or as muscle anchorage. Some species, notably incirrate octopuses, have lost their internal shell altogether, while in some it has been replaced by a chitinous support structure.
Belemnoids are an extinct group of marine cephalopod, very similar in many ways to the modern squid. Like them, the belemnoids possessed an ink sac, but, unlike the squid, they possessed ten arms of roughly equal length, and no tentacles. The name "belemnoid" comes from the Greek word βέλεμνον, belemnon meaning "a dart or arrow" and the Greek word είδος, eidos meaning "form".
Nautiloids are a group of marine cephalopods (Mollusca) which originated in the Late Cambrian and are represented today by the living Nautilus and Allonautilus. Fossil nautiloids are diverse and species rich, with over 2,500 recorded species. They flourished during the early Paleozoic era, when they constituted the main predatory animals. Early in their evolution, nautiloids developed an extraordinary diversity of shell shapes, including coiled morphologies and giant straight-shelled forms (orthocones). No orthoconic and only a handful of coiled species, the nautiluses, survive to the present day.
Aulacocerida is an order of primitive coleoid cephalopods, possibly derived from michelinoceraitids (Orthocerida) early in the Devonian, which in turn gave rise to the Belemnites.
Trachyteuthis is a genus of extinct octopodiform cephalopods, comprising six species: T. hastiformis, T. latipinnis, T. nusplingensis, T. teudopsiformis, T. covacevichi and T. chilensis.
Proteroctopus is an extinct genus of cephalopod that lived in the Middle Jurassic, approximately 164 million years ago. It is only known from a single species P. ribeti. The single fossil specimen assigned to this species originates from the Lower Callovian of Voulte-sur-Rhône in France. It is currently on display at the Musée de Paléontologie de La Voulte-sur-Rhône. While originally interpreted as an early octopus, a 2016 restudy of the specimen considered it to be a basal member of the Vampyropoda, less closely related to octopus or vampire squid than either of the two groups are to each other. A phylogenetic analysis by Kruta et. al indicates that Proteroctopus may be more closely related to the Vampyromorpha based on its unique morphology: two fins, head fused to the body, eight arms, two rows of oblique sucker, a gladius and absence of an ink sac. A 2022 phylogenetic analysis also found it to be more closely related to vampire squid than to octopuses.
Palaeoctopus is an extinct genus of octopuses that lived during the Late Cretaceous. It contains one valid species, P. newboldi, which has been found in Lebanon.
Boletzkyida is a primitive order of teuthid coleoid cephalopod: the boletzkyids are thought to be the earliest forms of coleoid cephalopods, and appear to form a link between nautiloid orthocerids and more advanced coleoids. Boletzkyida was named and described by Bandel, Reitner, and Sturmer in 1983 (B.R.&S) from specimens found in the Lower Devonian black slate in Germany.
Belemnitella is a genus of belemnite from the Late Cretaceous of Europe and North America.
Phragmoteuthis is a genus of extinct coleoid cephalopod known from the late Triassic to the lower Jurassic. Its soft tissue has been preserved; some specimens contain intact ink sacs. It had an internal phragmocone and ten arms.
Acanthoteuthis is a belemnite genus, a squid-like cephalopod with an internal shell from the Late Jurassic Epoch, related to modern coleoids.
Teudopsis is a genus of the coleoid cephalopod family Teudopsidae, known only from Lower Jurassic gladii. It has been reported from Alberta, Canada and Europe. During the Early Torcian period, the T. bollensis, T. bunelii, and T. subcostata all inhabited European seas. The differences in the anterior median field differentiates the three species. The life appearance of Teudopsis is probably best inferred by its close relative Trachyteuthis. A fossil with soft tissue preservation shows that this animal may have had two mantle fins, which may also be the case for Teudopsis.
Phragmoteuthida is an order of extinct coleoid cephalopods characterized by a fan-like teuthoid pro-ostracum attached to a belemnoid-like phragmocone.
Hematitida is a group of coleoid cephalopods known from the early Carboniferous Period. They are the oldest definite coleoids, although there are controversial claims for even older coleoids from the Devonian. Fossil hematitidans have so far been found only in Arkansas and Utah of the United States. The only family described so far is Hematitidae.
Belemnitida is an extinct order of squid-like cephalopods that existed from the Late Triassic to Late Cretaceous. Unlike squid, belemnites had an internal skeleton that made up the cone. The parts are, from the arms-most to the tip: the tongue-shaped pro-ostracum, the conical phragmocone, and the pointy guard. The calcitic guard is the most common belemnite remain. Belemnites, in life, are thought to have had 10 hooked arms and a pair of fins on the guard. The chitinous hooks were usually no bigger than 5 mm (0.20 in), though a belemnite could have had between 100 and 800 hooks in total, using them to stab and hold onto prey.
All extant cephalopods have a two-part beak, or rostrum, situated in the buccal mass and surrounded by the muscular head appendages. The dorsal (upper) mandible fits into the ventral (lower) mandible and together they function in a scissor-like fashion. The beak may also be referred to as the mandibles or jaws. These beaks are different from bird beaks because they crush bone while most bird beaks do not.
Cephalopod egg fossils are the fossilized remains of eggs laid by cephalopods. The fossil record of cephalopod eggs is scant since their soft, gelatinous eggs decompose quickly and have little chance to fossilize. Eggs laid by ammonoids are the best known and only a few putative examples of these have been discovered. The best preserved of these were discovered in the Jurassic Kimmeridge Clay of England. Currently no belemnoid egg fossils have ever been discovered although this may be because scientists have not properly searched for them rather than an actual absence from the fossil record.
Muensterella is a fossil stem-octopod known from a handful of specimens from the Late Jurassic lithographic limestones of southern Germany and the upper Cretaceous of Germany and Texas. Another indeterminate specimen of the Genus is known from the Tithonian Nordenskjöld Formation of Antarctica.
Clarkeiteuthis is a genus of extinct belemnoid cephalopod known from the lower Jurassic in Germany and England. Described two species, C. conocauda and C. montefiorei are originally described as species of phragmoteuthid Phragmoteuthis, but got their own genus and moved to Diplobelida.
Antarcticeras is an extinct genus of enigmatic cephalopod from the Eocene of Antarctica. It contains a single species, A.nordenskjoeldi. It is either considered the last of the "orthocone"-type cephalopods, the only member of its subclass Paracoleoidea & a descendant of the orthoceratids, and a remarkable example of convergent evolution with coleoid cephalopods, or an oegospid squid and a transitional form in the development of the modern squid gladius, of which it is the only preserved example.
1 2 3 4 Dirk Fuchs; Helmut Keupp; Vasilij Mitta & Theo Engeser (2007). "Ultrastructural Analyses on the Conotheca of the genus Belemnotheutis (Belemnitida: Coleoidea)". In Neil H. Landman; Richard Arnold Davis & Royal H. Mapes (eds.). Cephalopods present and past: new insights and fresh perspectives. Springer. ISBN978-1-4020-6461-6.
1 2 3 4 5 6 7 Jeletzky, J.A. 1966. Comparative Morphology, Phylogeny, and Classification of Fossil Coleoidea; Mollusca pp 1–162; The University of Kansas, Paleontological Contributions
1 2 3 Philip R. Wilby et al 2008. Preserving the unpreservable: a lost world rediscovered at Christian Malford, UK. Geology Today Vol 24(3). Blackwell Publishing Ltd.
↑ Jack Sepkoski 2002. A compendium of fossil marine animal genera (Cephalopoda entry). Bulletins of American Paleontology, 364 p.560
↑ Engeser, T.S. and J. Reitner. 1992. Ein neues Exemplar von Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) aus dem Solnhofener Plattenkalk (Untertithonium) von Wintershof, Bayern. Archaeopteryx 10:13-17.
↑ Woodward, Horace Bolingbroke. The history of the Geological Society of London Geological Society, London 1978, page 115
↑ Pharaoh, J.B.1837. Fossil Remains of naked Mollusks, Pens, and Ink-Bags of Loligo. Madras Journal of Literature and Science vol5, issue 14. pp 403–406. Madras Literary Society, Auxiliary Royal Asiatic Society |
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.