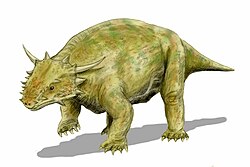
Pareiasaurs (meaning "cheek lizards") are an extinct clade of large, herbivorous reptiles (traditionally considered "parareptiles") that existed during the Permian period. Members of the group were armoured with osteoderms which covered large areas of the body. They first appeared in southern Pangea during the Middle Permian, before becoming globally distributed during the Late Permian. Pareiasaurs were the largest reptiles of the Permian, some reaching sizes over 1 tonne (2,200lb), equivalent to the largest contemporary therapsids. Pareiasaurs became extinct in the end-Permian mass extinction event.
Restoration of BradysaurusSize of Scutosaurus, one of the largest pareiasaurs, compared to a human
Pareiasaurs ranged in size from 60 to 300 centimetres (2.0 to 9.8ft) long, with the largest pareiasaurs like Scutosaurus and Bradysaurus estimated to exceed 1,000 kilograms (2,200lb) in body mass.[1][2] The limbs of many parieasaurs were extremely robust, likely to account for the increased stress on their limbs caused by their typically sprawling posture.[1][2] The cow-sized Bunostegos differed from other pareiasaurs by having a more upright limb posture, being amongst the first amniotes to develop this trait.[3] Pareiasaurs were protected by bony scutes called osteoderms that were set into the skin.[4] Their skulls were heavily ornamented with bosses, rugose ridges, and bumps.[5] Their leaf-shaped multi-cusped teeth resemble those of iguanas, indicating a herbivorous diet.[6] The body probably housed an extensive digestive tract.[1] Most authors have assumed a terrestrial lifestyle for pareiasaurs. A 2008 bone microanatomy study suggested a more aquatic, plausibly amphibious lifestyle,[7] but a later 2019 study found that the bone histology provided no direct evidence of this lifestyle.[8]
Evolutionary history
Among reptilian groups, pareiasaurs are thought to be most closely related to the nycteroleterids as part of the clade Pareiasauromorpha.[9] Pareiasaurs filled the large herbivore niche (or guild) that had been occupied early in the Permian period by the caseid pelycosaurs and, before them, the diadectid reptiliomorphs.[8] They are much larger than the diadectids, more similar to the giant caseid pelycosaur Cotylorhynchus. Although the last Pareiasaurs were no larger than the first types (indeed, many of the last ones became smaller), there was a definite tendency towards increased armour as the group developed. Pareiasaurs first appeared in the fossil record in the Middle Permian (Guadalupian) of Southern Pangaea, before dispersing into Northern Pangaea and gaining a cosmopolitan distribution during the Late Permian (Lopingian).[10]
Classification
Some paleontologists considered that pareiasaurs were direct ancestors of modern turtles. Pareiasaur skulls have several turtle-like features, and in some species the scutes have developed into bony plates, possibly the precursors of a turtle shell.[11] Jalil and Janvier, in a large analysis of pareiasaur relationships, also found turtles to be close relatives of the "dwarf" pareiasaurs, such as Pumiliopareia.[12] However, the discovery of Pappochelys argues against a potential pareisaurian relationship to turtles,[13] and DNA evidence indicates that living turtles are more closely related to living archosaurs than lepidosaurs, and therefore cladistically diapsids.[14]
Pareiasaurs are currently generally considered to be an earlier branch of the reptilian family tree that diverged before the common ancestor of living reptiles including turtles. Pareiasaurs were long historically included with many other primitive reptiles in the group "Parareptilia", but recent research has questioned the validity of this group.[15]
Associated clades
Hallucicrania (Lee 1995): This clade was coined by MSY Lee for Lanthanosuchidae + (Pareiasauridae + Testudines). Lee's pareiasaur hypothesis has become untenable due to the diapsid features of the stem turtle Pappochelys and the potential testudinatan nature of Eunotosaurus. Recent cladistic analyses reveal that lanthanosuchids have a much more basal position in the Procolophonomorpha, and that the nearest sister taxon to the pareiasaurs are the rather unexceptional and conventional looking nycteroleterids (Müller & Tsuji 2007, Lyson et al. 2010) the two being united in the clade Pareiasauromorpha (Tsuji et al. 2012).
Pareiasauroidea (Nopcsa, 1928): This clade (as opposed to the superfamily or suborder Pareiasauroidea) was used by Lee (1995) for Pareiasauridae + Sclerosaurus. More recent cladistic studies place Sclerosaurus in the procolophonid subfamily Leptopleuroninae (Cisneros 2006, Sues & Reisz 2008), which means the similarities with pareiasaurs are the result of convergences.
Pareiasauria (Seeley, 1988): If neither Lanthanosuchidae or Testudines are included in the clade, the Pareiasauria only contains the monophyletic family Pareiasauridae.
Phylogeny
Cladogram of Reptilia after Jenkins et al 2025, with orange bars highlighting taxa formerly considered parareptiles:[15]
In 2025, Jian Yi and Jun Liu described Yinshanosaurus as a new Chinese pareiasaur based on two well-preserved, nearly complete skulls and an incomplete, partially articulated skeleton. Their publication included a phylogenetic analysis of pareiasaurs, the results of which are displayed in the cladogram below:[16]
↑Scheyer, T. M. & Sander, P. M. (2009). "Bone microstructures and mode of skeletogenesis in osteoderms of three pareiasaur taxa from the Permian of South Africa". Journal of Evolutionary Biology. 22 (6): 1153–1162. doi:10.1111/j.1420-9101.2009.01732.x. PMID19416416.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Van Den Brandt, Marc Johan; Abdala, Fernando; Rubidge, Bruce Sidney (2019). "Cranial morphology and phylogenetic relationships of the Middle Permian pareiasaur Embrithosaurus schwarzi from the Karoo Basin of South Africa". Zoological Journal of the Linnean Society. doi:10.1093/zoolinnean/zlz064.
↑LEE, M. S. Y. (1995). "Historical burden in systematics and the interrelationships of 'parareptiles'". Biological Reviews of the Cambridge Philosophical Society. 70 (3): 459–547. doi:10.1111/j.1469-185x.1995.tb01197.x. S2CID85790423.
↑Crawford, Nicholas G.; Parham, James F.; Sellas, Anna B.; Faircloth, Brant C.; Glenn, Travis C.; Papenfuss, Theodore J.; Henderson, James B.; Hansen, Madison H.; Simison, W. Brian (February 2015). "A phylogenomic analysis of turtles". Molecular Phylogenetics and Evolution. 83: 250–257. doi:10.1016/j.ympev.2014.10.021. PMID25450099. Archived from the original on 2024-04-15. Retrieved 2021-05-19.
↑Yi, Jian; Liu, Jun (2025-06-04). "The tetrapod fauna of the upper Permian Naobaogou Formation of China: a new mid-sized pareiasaur Yinshanosaurus angustus and its implications for the phylogenetic relationships of pareiasaurs". Papers in Palaeontology. 11 (3). doi:10.1002/spp2.70020. ISSN2056-2799.
Further reading
Carroll, R. L., (1988), Vertebrate Paleontology and Evolution, W.H. Freeman & Co. New York, p.205
Kuhn, O, 1969, Cotylosauria, part 6 of Handbuch der Palaoherpetologie (Encyclopedia of Palaeoherpetology), Gustav Fischer Verlag, Stuttgart & Portland
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.