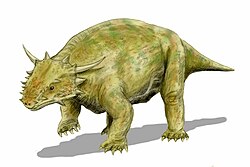
Bradysaurus is a genus of large, primitive and widespread pareiasaur. They possessed a covering of armoured scutes, likely serving as defense against their main predators, the gorgonopsians.
Bradysaurus was 2.5–3m (8ft 2in– 9ft 10in) in length[1] and half a tonne to a tonne in weight. The skull was large (about 42 to 48centimeters long), broad and rounded at the front. It was coarsely sculptured and knobby, with the sutures between the bones not clearly visible.
The marginal teeth were high-crowned, with only a few cusps, which is a primitive characteristic. The feet were short and broad, the phalangeal count being 2,3,3,3,2 on the fore-foot and 2,3,3,4,3 on the hind. The whole body is protected by dermal scutes, although these are not as thick or heavy as in more advanced forms.
Classification and species
Bradysaurus is the only member of the subfamily Bradysaurinae. It is the most primitive known pareiasaur and can be considered a good ancestral type from which the others developed. Its large dimensions show that, even very early in their evolutionary history, these strange animals had already attained an optimal size. Even later, more advanced forms, like Scutosaurus, were no larger. The advantage of large size was to provide defense against predators and to maintain a stable body temperature (gigantothermy).
Kuhn 1969 lists no fewer than nine species for this genus, but this is certainly an excessive number.[2] Boonstra 1969 distinguishes only four species on the basis of tooth structure,[3] two of which Kuhn places in the genus Embrithosaurus.[2] The genera Brachypareia, Bradysuchus, Koalemasaurus, and Platyoropha are synonyms of Bradysaurus.
B. baini
B. baini (Seeley, 1892) is from the Tapinocephalus zone, Lower Beaufort Beds, Karoo basin, South Africa. This is the type species for the genus. The quadra-jugal region (cheek-bones) were only moderately developed. The snout was broad and rounded and there were 15 or 16pairs of overlapping teeth in each jaw. This animal could be considered a generic early pareiasaur. According to Lee, 1997, the available material of B. baini lacks distinguishing autapomorphies or characteristics.[4]
B. Seeleyi
B. seeleyi (Haughton and Boonstra, 1929) is from the Tapinocephalus zone, Lower Beaufort Beds, Karoo basin, South Africa. This is a less common form. Boonstra, 1969, considered this a valid species of Bradysaurus[3] and Lee, 1997,[4] considers this animal a sister group to more advanced pareiasaurs. B. seelyi seems to be closely related to Nochelesaurus and Embrithosaurus. In contrast to the more numerous but similarly sized B. baini, the cheekbones were heavy and greatly enlarged. There were 19 or 20 pairs of strongly overlapping teeth on each jaw.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.