Silicon is a chemical element; it has symbol Si and atomic number 14. It is a hard, brittle crystalline solid with a blue-grey metallic luster, and is a tetravalent metalloid and semiconductor. It is a member of group 14 in the periodic table: carbon is above it; and germanium, tin, lead, and flerovium are below it. It is relatively unreactive. Silicon is a significant element that is essential for several physiological and metabolic processes in plants. Silicon is widely regarded as the predominant semiconductor material due to its versatile applications in various electrical devices such as transistors, solar cells, integrated circuits, and others. These may be due to its significant band gap, expansive optical transmission range, extensive absorption spectrum, surface roughening, and effective anti-reflection coating.

A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

Chert is a hard, fine-grained sedimentary rock composed of microcrystalline or cryptocrystalline quartz, the mineral form of silicon dioxide (SiO2). Chert is characteristically of biological origin, but may also occur inorganically as a chemical precipitate or a diagenetic replacement, as in petrified wood.

The biological pump (or ocean carbon biological pump or marine biological carbon pump) is the ocean's biologically driven sequestration of carbon from the atmosphere and land runoff to the ocean interior and seafloor sediments. In other words, it is a biologically mediated process which results in the sequestering of carbon in the deep ocean away from the atmosphere and the land. The biological pump is the biological component of the "marine carbon pump" which contains both a physical and biological component. It is the part of the broader oceanic carbon cycle responsible for the cycling of organic matter formed mainly by phytoplankton during photosynthesis (soft-tissue pump), as well as the cycling of calcium carbonate (CaCO3) formed into shells by certain organisms such as plankton and mollusks (carbonate pump).
High-nutrient, low-chlorophyll (HNLC) regions are regions of the ocean where the abundance of phytoplankton is low and fairly constant despite the availability of macronutrients. Phytoplankton rely on a suite of nutrients for cellular function. Macronutrients are generally available in higher quantities in surface ocean waters, and are the typical components of common garden fertilizers. Micronutrients are generally available in lower quantities and include trace metals. Macronutrients are typically available in millimolar concentrations, while micronutrients are generally available in micro- to nanomolar concentrations. In general, nitrogen tends to be a limiting ocean nutrient, but in HNLC regions it is never significantly depleted. Instead, these regions tend to be limited by low concentrations of metabolizable iron. Iron is a critical phytoplankton micronutrient necessary for enzyme catalysis and electron transport.

Orthosilicic acid is an inorganic compound with the formula Si(OH)4. Although rarely observed, it is the key compound of silica and silicates and the precursor to other silicic acids [H2xSiOx+2]n. Silicic acids play important roles in biomineralization and technology. It is the parent acid of the orthosilicate anion SiO4−
4.
Lithogenic silica (LSi) is silica (SiO2) derived from terrigenous rock (Igneous, metamorphic, and sedimentary), lithogenic sediments composed of the detritus of pre-existing rock, volcanic ejecta, extraterrestrial material, and minerals such silicate. Silica is the most abundant compound in the Earth's crust (59%) and the main component of almost every rock (>95%).
Biogenic silica (bSi), also referred to as opal, biogenic opal, or amorphous opaline silica, forms one of the most widespread biogenic minerals. For example, microscopic particles of silica called phytoliths can be found in grasses and other plants.

Marine sediment, or ocean sediment, or seafloor sediment, are deposits of insoluble particles that have accumulated on the seafloor. These particles either have their origins in soil and rocks and have been transported from the land to the sea, mainly by rivers but also by dust carried by wind and by the flow of glaciers into the sea, or they are biogenic deposits from marine organisms or from chemical precipitation in seawater, as well as from underwater volcanoes and meteorite debris.

Siliceous ooze is a type of biogenic pelagic sediment located on the deep ocean floor. Siliceous oozes are the least common of the deep sea sediments, and make up approximately 15% of the ocean floor. Oozes are defined as sediments which contain at least 30% skeletal remains of pelagic microorganisms. Siliceous oozes are largely composed of the silica based skeletons of microscopic marine organisms such as diatoms and radiolarians. Other components of siliceous oozes near continental margins may include terrestrially derived silica particles and sponge spicules. Siliceous oozes are composed of skeletons made from opal silica SiO2·nH2O, as opposed to calcareous oozes, which are made from skeletons of calcium carbonate (CaCO3·nH2O) organisms (i.e. coccolithophores). Silica (Si) is a bioessential element and is efficiently recycled in the marine environment through the silica cycle. Distance from land masses, water depth and ocean fertility are all factors that affect the opal silica content in seawater and the presence of siliceous oozes.

The carbonate–silicate geochemical cycle, also known as the inorganic carbon cycle, describes the long-term transformation of silicate rocks to carbonate rocks by weathering and sedimentation, and the transformation of carbonate rocks back into silicate rocks by metamorphism and volcanism. Carbon dioxide is removed from the atmosphere during burial of weathered minerals and returned to the atmosphere through volcanism. On million-year time scales, the carbonate-silicate cycle is a key factor in controlling Earth's climate because it regulates carbon dioxide levels and therefore global temperature.

Fragilariopsis kerguelensis, is a pennate diatom native to the Southern Ocean. It has been characterized as "the most abundant diatom in the Antarctic Seas".
Reverse weathering generally refers to a process of clay neoformation consuming cations and alkalinity in a way unrelated to the weathering of silicates. More specifically reverse weathering refers to the formation of authigenic clay minerals from the reaction of 1) biogenic silica with aqueous cations or cation-bearing oxides or 2) cation poor precursor clays with dissolved cations or cation-bearing oxides.

Marine biogeochemical cycles are biogeochemical cycles that occur within marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. These biogeochemical cycles are the pathways chemical substances and elements move through within the marine environment. In addition, substances and elements can be imported into or exported from the marine environment. These imports and exports can occur as exchanges with the atmosphere above, the ocean floor below, or as runoff from the land.

Aluminum is the third most abundant element in the lithosphere at 82,000 ppm. It occurs in low levels, 0.9 ppm, in humans. Aluminum is known to be an ecotoxicant and expected to be a health risk to people. Global primary production (GPP) of aluminum was about 52 million tons in 2013 and remains one of the world's most important metals. It is used for infrastructure, vehicles, aviation, energy and more due to its lightweight, ductility, and cheap cost. Aluminum is harvested from gibbsite, boehmite, and diaspore which make up bauxite. The aluminum cycle is the biogeochemical cycle by which aluminum is moved through the environment by natural and anthropogenic processes. The biogeochemical cycle of aluminum is integral with silicon and phosphorus. For example, phosphates store aluminum that has been sedimented and aluminum is found in diatoms. Aluminum has been found to prevent growth in organisms by making phosphates less available. The humans/lithosphere ratio (B/L) is very low at 0.000011. This level shows that aluminum is more essential in the lithospheric cycle than in the biotic cycle.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.

Particulate inorganic carbon (PIC) can be contrasted with dissolved inorganic carbon (DIC), the other form of inorganic carbon found in the ocean. These distinctions are important in chemical oceanography. Particulate inorganic carbon is sometimes called suspended inorganic carbon. In operational terms, it is defined as the inorganic carbon in particulate form that is too large to pass through the filter used to separate dissolved inorganic carbon.
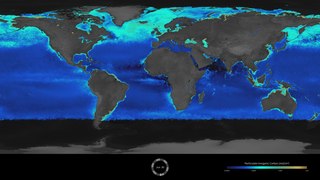
The Great Calcite Belt (GCB) refers to a region of the ocean where there are high concentrations of calcite, a mineral form of calcium carbonate. The belt extends over a large area of the Southern Ocean surrounding Antarctica. The calcite in the Great Calcite Belt is formed by tiny marine organisms called coccolithophores, which build their shells out of calcium carbonate. When these organisms die, their shells sink to the bottom of the ocean, and over time, they accumulate to form a thick layer of calcite sediment.

In geology, silicification is a petrification process in which silica-rich fluids seep into the voids of Earth materials, e.g., rocks, wood, bones, shells, and replace the original materials with silica (SiO2). Silica is a naturally existing and abundant compound found in organic and inorganic materials, including Earth's crust and mantle. There are a variety of silicification mechanisms. In silicification of wood, silica permeates into and occupies cracks and voids in wood such as vessels and cell walls. The original organic matter is retained throughout the process and will gradually decay through time. In the silicification of carbonates, silica replaces carbonates by the same volume. Replacement is accomplished through the dissolution of original rock minerals and the precipitation of silica. This leads to a removal of original materials out of the system. Depending on the structures and composition of the original rock, silica might replace only specific mineral components of the rock. Silicic acid (H4SiO4) in the silica-enriched fluids forms lenticular, nodular, fibrous, or aggregated quartz, opal, or chalcedony that grows within the rock. Silicification happens when rocks or organic materials are in contact with silica-rich surface water, buried under sediments and susceptible to groundwater flow, or buried under volcanic ashes. Silicification is often associated with hydrothermal processes. Temperature for silicification ranges in various conditions: in burial or surface water conditions, temperature for silicification can be around 25°−50°; whereas temperatures for siliceous fluid inclusions can be up to 150°−190°. Silicification could occur during a syn-depositional or a post-depositional stage, commonly along layers marking changes in sedimentation such as unconformities or bedding planes.
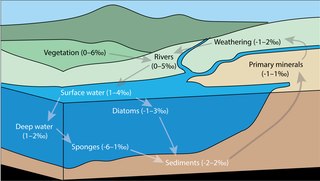
Silicon isotope biogeochemistry is the study of environmental processes using the relative abundance of Si isotopes. As the relative abundance of Si stable isotopes varies among different natural materials, the differences in abundance can be used to trace the source of Si, and to study biological, geological, and chemical processes. The study of stable isotope biogeochemistry of Si aims to quantify the different Si fluxes in the global biogeochemical silicon cycle, to understand the role of biogenic silica within the global Si cycle, and to investigate the applications and limitations of the sedimentary Si record as an environmental and palaeoceanographic proxy.