
The major histocompatibility complex (MHC) is a large locus on vertebrate DNA containing a set of closely linked polymorphic genes that code for cell surface proteins essential for the adaptive immune system. These cell surface proteins are called MHC molecules.

An antigen-presenting cell (APC) or accessory cell is a cell that displays antigen bound by major histocompatibility complex (MHC) proteins on its surface; this process is known as antigen presentation. T cells may recognize these complexes using their T cell receptors (TCRs). APCs process antigens and present them to T-cells.

MHC class I molecules are one of two primary classes of major histocompatibility complex (MHC) molecules and are found on the cell surface of all nucleated cells in the bodies of vertebrates. They also occur on platelets, but not on red blood cells. Their function is to display peptide fragments of proteins from within the cell to cytotoxic T cells; this will trigger an immediate response from the immune system against a particular non-self antigen displayed with the help of an MHC class I protein. Because MHC class I molecules present peptides derived from cytosolic proteins, the pathway of MHC class I presentation is often called cytosolic or endogenous pathway.

HLA-DR is an MHC class II cell surface receptor encoded by the human leukocyte antigen complex on chromosome 6 region 6p21.31. The complex of HLA-DR and peptide, generally between 9 and 30 amino acids in length, constitutes a ligand for the T-cell receptor (TCR). HLA were originally defined as cell surface antigens that mediate graft-versus-host disease. Identification of these antigens has led to greater success and longevity in organ transplant.

HLA class II histocompatibility antigen, DR alpha chain is a protein that in humans is encoded by the HLA-DRA gene. HLA-DRA encodes the alpha subunit of HLA-DR. Unlike the alpha chains of other Human MHC class II molecules, the alpha subunit is practically invariable. However it can pair with, in any individual, the beta chain from 3 different DR beta loci, DRB1, and two of any DRB3, DRB4, or DRB5 alleles. Thus there is the potential that any given individual can form 4 different HLA-DR isoforms.
Antigen presentation is a vital immune process that is essential for T cell immune response triggering. Because T cells recognize only fragmented antigens displayed on cell surfaces, antigen processing must occur before the antigen fragment can be recognized by a T-cell receptor. Specifically, the fragment, bound to the major histocompatibility complex (MHC), is transported to the surface of the cell, a process known as presentation. If there has been an infection with viruses or bacteria, the cell will present an endogenous or exogenous peptide fragment derived from the antigen by MHC molecules. There are two types of MHC molecules which differ in the behaviour of the antigens: MHC class I molecules (MHC-I) bind peptides from the cell cytosol, while peptides generated in the endocytic vesicles after internalisation are bound to MHC class II (MHC-II). Cellular membranes separate these two cellular environments - intracellular and extracellular. Each T cell can only recognize tens to hundreds of copies of a unique sequence of a single peptide among thousands of other peptides presented on the same cell, because an MHC molecule in one cell can bind to quite a large range of peptides. Predicting which antigens will be presented to the immune system by a certain MHC/HLA type is difficult, but the technology involved is improving.

MHC Class II molecules are a class of major histocompatibility complex (MHC) molecules normally found only on professional antigen-presenting cells such as dendritic cells, mononuclear phagocytes, some endothelial cells, thymic epithelial cells, and B cells. These cells are important in initiating immune responses.
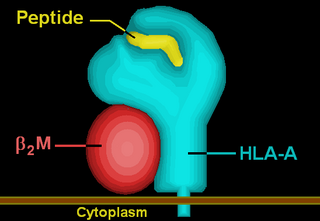
HLA-A is a group of human leukocyte antigens (HLA) that are encoded by the HLA-A locus, which is located at human chromosome 6p21.3. HLA is a major histocompatibility complex (MHC) antigen specific to humans. HLA-A is one of three major types of human MHC class I transmembrane proteins. The others are HLA-B and HLA-C. The protein is a heterodimer, and is composed of a heavy α chain and smaller β chain. The α chain is encoded by a variant HLA-A gene, and the β chain (β2-microglobulin) is an invariant β2 microglobulin molecule. The β2 microglobulin protein is encoded by the B2M gene, which is located at chromosome 15q21.1 in humans.

HLA class I histocompatibility antigen, alpha chain E (HLA-E) also known as MHC class I antigen E is a protein that in humans is encoded by the HLA-E gene. The human HLA-E is a non-classical MHC class I molecule that is characterized by a limited polymorphism and a lower cell surface expression than its classical paralogues. The functional homolog in mice is called Qa-1b, officially known as H2-T23.

CD94, also known as killer cell lectin-like receptor subfamily D, member 1 (KLRD1) is a human gene.

Minor histocompatibility antigen are peptides presented on the cellular surface of donated organs that are known to give an immunological response in some organ transplants. They cause problems of rejection less frequently than those of the major histocompatibility complex (MHC). Minor histocompatibility antigens (MiHAs) are diverse, short segments of proteins and are referred to as peptides. These peptides are normally around 9-12 amino acids in length and are bound to both the major histocompatibility complex (MHC) class I and class II proteins. Peptide sequences can differ among individuals and these differences arise from SNPs in the coding region of genes, gene deletions, frameshift mutations, or insertions. About a third of the characterized MiHAs come from the Y chromosome. Prior to becoming a short peptide sequence, the proteins expressed by these polymorphic or diverse genes need to be digested in the proteasome into shorter peptides. These endogenous or self peptides are then transported into the endoplasmic reticulum with a peptide transporter pump called TAP where they encounter and bind to the MHC class I molecule. This contrasts with MHC class II molecules's antigens which are peptides derived from phagocytosis/endocytosis and molecular degradation of non-self entities' proteins, usually by antigen-presenting cells. MiHA antigens are either ubiquitously expressed in most tissue like skin and intestines or restrictively expressed in the immune cells.

HLA-DM is an intracellular protein involved in the mechanism of antigen presentation on antigen presenting cells (APCs) of the immune system. It does this by assisting in peptide loading of major histocompatibility complex (MHC) class II membrane-bound proteins. HLA-DM is encoded by the genes HLA-DMA and HLA-DMB.
Major histocompatibility complex, class II, DR beta 4, also known as HLA-DRB4, is a human gene.

HLA-G histocompatibility antigen, class I, G, also known as human leukocyte antigen G (HLA-G), is a protein that in humans is encoded by the HLA-G gene.
HLA class II histocompatibility antigen gamma chain also known as HLA-DR antigens-associated invariant chain or CD74, is a protein that in humans is encoded by the CD74 gene. The invariant chain is a polypeptide which plays a critical role in antigen presentation. It is involved in the formation and transport of MHC class II peptide complexes for the generation of CD4+ T cell responses. The cell surface form of the invariant chain is known as CD74. CD74 is a cell surface receptor for the cytokine macrophage migration inhibitory factor (MIF).
MHC class I polypeptide–related sequence A (MICA) is a highly polymorphic cell surface glycoprotein encoded by the MICA gene located within MHC locus. MICA is related to MHC class I and it has similar domain structure, however, it is not associated with β2-microglobulin nor binds peptides as conventional MHC class I molecules do. MICA rather functions as a stress-induced ligand (as a danger signal) for integral membrane protein receptor NKG2D ("natural-killer group 2, member D"). MICA is broadly recognized by NK cells, γδ T cells, and CD8+ αβ T cells which carry NKG2D receptor on their cell surface and which are activated via this interaction.

HLA class II histocompatibility antigen, DRB3-1 beta chain is a protein that in humans is encoded by the HLA-DRB3 gene.
HLA class II histocompatibility antigen, DM beta chain is a protein that in humans is encoded by the HLA-DMB gene.

HLA class II histocompatibility antigen, DM alpha chain is a protein that in humans is encoded by the HLA-DMA gene.
HLA class II histocompatibility antigen, DO beta chain is a protein that in humans is encoded by the HLA-DOB gene.



