Palaeontinidae, commonly known as giant cicadas, is an extinctfamily of cicadomorphs. They existed from the Late Triassic to the Early Cretaceous. The family contains around 30 to 40 genera and around a hundred species.[1] They are thought to have had a similar ecology to modern cicadas as feeders on plant xylem fluids. Despite being described as "giant cicadas"(with the wingspan of some species exceeding 15 centimetres (5.9in)[2]),[3] they are not particularly closely related to true cicadas.[4]
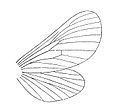
Palaeontinids had large bodies covered with bristles (setae). They had small heads and broad wings. They superficially resemble moths.[7][8] Large palaeontinids like Colossocossus had forewings that reached the length of 57 to 71mm (2.2 to 2.8in).[9] They possessed an inflated frons and a long rostrum (piercing and sucking mouthpart), indicating that they fed on xylem fluids like some other modern hemipterans,[10] including living cicadas to which they have often been compared.[11]
Some authors have proposed that the host plants of palaeontinids to be ginkgophytes based on the geographic distribution of both groups, however other authors have argued that this association is likely to be spurious, given that paleontinids also occur in areas with no ginkgophytes. Some authors have suggested that the decline of gymnosperms and the rise of angiosperms (flowering plants) during the Cretaceous could have been a factor in their extinction.[9] Numerous newly evolved insectivorous animals (feathered theropods, primitive mammals, and early birds) may have also contributed significantly to their extinction.[12]
Most species of palaeontinids exhibit cryptic coloration.[13] The patterns on their wings protected them as they perched on branches and fed on sap. They may also have served as secondary sexual characteristics. The color patterns can vary slightly within the same species.[10]
Illustration of a palaeontinid being pursued by a primitive bird during the Early Cretaceous
Palaeontinids, like modern cicadas, possess four membranous wings supported by veins. The length and width ratio of the wings can vary within the same species, sometimes as a result of fossil preservation.[10] Early Jurassic palaeontinids, like Suljuktocossus, exhibit the most primitive wing forms in the family.[14] The forewing was elliptical with the "nodal line" (the area where the wing bends during flight, also known as the "transverse flexion line") more or less dissecting through the center of the wing. The hindwing was short and broad. The bases of the forewings overlapped that of the hindwings like in modern butterflies. Taken together with their large bodies, these characteristics indicate that they were fast but moderately versatile fliers.[15]
In contrast, later palaeontinids like the Upper Jurassic Eocicada and Early Cretaceous Ilerdocossus had triangular forewings with the flexion line closer to the base. They had smaller and narrower hindwings that did not overlap with the forewing. These indicate that they were highly versatile fliers, able to fly with a wide range of speeds and agility like modern wasps and sphinx moths.[15] They also possessed changes to the leading edge of their forewings, suggesting an overall gain in lift.[14] The different wing shape of later palaeontinids may have evolved to more effectively escape from flying predators like early birds.[11]
The trend of forewing elongation is most evident in members of the family Mesogereonidae, an early offshoot and close relatives of palaeontinids.[16]
The family was first erected by the Austrian entomologist Anton Handlirsch in 1908. Like Butler, Handlirsch insisted that palaeontinids were members of lepidopteranHeteroneura (butterflies and moths). Palaeontinids were then only known mostly from poorly preserved specimens like Palaeontina and Eocicada. He claimed they were related to the extant family Limacodidae (slug moths).[17] The English entomologist Edward Meyrick supported the lepidopteran conclusion, though he believed they belonged to the family Hepialidae (ghost moths) instead. He said "There is little doubt that it [i.e. Palaeontina oolitica] belongs to the Hepialidae."[5]
The Belgian entomologist Auguste Lameere challenged this conclusion, claiming palaeontinids were more closely related to the extant family Cicadidae (cicadas). The English-Australian entomologist and geologistRobert John Tillyard supported Lameere's conclusion, noting that the wings of palaeontinid fossils lacked the characteristic scales of lepidopterans but instead had tubercules, pits, and cross-ridges like those found in modern cicadas.[17] He also cited characteristics of wing venation that distinctly differs from that of lepidopterans.[5]
The name Cicadomorphidae was once proposed as a replacement for the name Palaeontinidae in 1956 by the Australian entomologist J.W. Evans. This was because of Handlirsch's earlier insistence that the type speciesPalaeontina oolitica may not have been Hemipteran. However, Evans later conceded that retaining the name Palaeontinidae was preferable as the drawings Handlirsch based his conclusions on were from badly preserved specimens.[20]
Evolution
Riek (1976) originally considered Palaeontinoidea to be the descendants of the family Cicadoprosbolidae (currently known as the family Tettigarctidae), insects believed to be transitional between the ancestral cicada-like family Prosbolidae and the modern family Cicadidae.[14]
Wang et al (2009), however, notes that palaeontinoids more closely resemble prosbolids in agreement with earlier studies by Wootton (1971), Shcherbakov (1984), and Shcherbakov and Popov (2002). They conclude that palaeontinoids descended directly from the family Prosbolidae rather than from tettigarctids.[14] Modern cicadas therefore, did not descend directly from Palaeontinidae.
Within Palaeontinoidea, the family Dunstaniidae (Upper Permian to Lower Jurassic of Australia, South Africa, and China) is ancestral to palaeontinids. Both are distinct from the only other member of the superfamily, the more primitive and specialized family Mesogereonidae (Upper Triassic of Australia and South Africa).[14]
The oldest known member of the group is Hallakkungisfrom South Korea dating to the Norian stage of the Late Triassic (ca. 227 – ca. 208.5 Mya)[21] and the youngest members are from the late Aptian age of the Lower Cretaceous (~115-113 Mya).[12][18] They achieved their greatest diversity during the Jurassic period.[22]
†PalaeontinodesMartynov, 1937 - (= IjacossusBecker-Migdisova, 1950; Shcherbakov 1985) Sulyukta Formation, Sagul Formation, Haifanggou Formation, Cheremkhovskaya Formation, Early to Middle Jurassic, Central and North Asia; Middle Jurassic, Central Asia
†ParawonnacottellaUeda, 1997 - Crato Formation, Early Cretaceous, Eastern South America
†PhragmatoecicossusBecker-Migdisova, 1949 - Sagul Formation, Early to Middle Jurassic, Central Asia
†PhragmatoecitesOppenheim, 1885 - Cheremkhovskaya Formation, Early to Middle Jurassic, North Asia
†PlachutellaBecker-Migdisova, 1949 - Badaowan Formation, Jiulongshan Formation, Daohugou, Karabastau Formation, Sagul Formation, Cheremkhovskaya Formation, Early to Late Jurassic, Central and Eastern Asia
†ProlystraOppenheim, 1888 syn Beloptesis- Solnhofen Formation, Late Jurassic, Central Europe
†ProtopsycheHandlirsch, 1906–1908 - Solnhofen Formation, Late Jurassic, Central Europe
†ShurabocossusBecker-Migdisova, 1949 - Sagul Formation, Early to Middle Jurassic, Central Asia
†SinopalaeocossusHong, 1983- (= QuadraticossusWang & Ren, 2007a) Daohugou, Haifanggou Formation, Middle Jurassic, East Asia
†SuljuktajaBecker-Migdisova, 1949 - Sulyukta Formation, Early to Middle Jurassic, Central Asia
†SuljuktocossusBecker-Migdisova, 1949 - (= FletcherianaEvans, 1956in partim) Sulyukta Formation, Early Jurassic, Central Asia; Daohugou Middle Jurassic, North Asia
†SynapocossusWang et al. 2013 Daohugou Middle Jurassic, East Asia
↑R. Wootton (1971). "The Evolution of Cicadoidea (Homoptera)"(PDF). Proceedings: XIII International Congress of Entomology, Moscow (1): 318–319. Retrieved July 15, 2011.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.