Neurotensin shares significant sequence similarity in its 6 C-terminalamino acids with several other neuropeptides, including neuromedin N (which is derived from the same precursor). This C-terminal region is responsible for the full biological activity, the N-terminal portion having a modulatory role. The neurotensin/neuromedin N precursor can also be processed to produce large 125–138 amino acidpeptides with the neurotensin or neuromedin N sequence at their C terminus. These large peptides appear to be less potent than their smaller counterparts, but are also less sensitive to degradation and may represent endogenous, long-lasting activators in a number of pathophysiological situations.
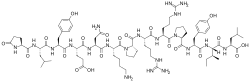
The sequence of bovine neurotensin was determined to be pyroGlu-Leu-Tyr-Glu-Asn-Lys-Pro-Arg-Arg-Pro-Tyr-Ile-Leu-OH.[7] Neurotensin is synthesized as part of a 169 or 170 amino acid precursor protein that also contains the related neuropeptide neuromedin N.[8][9] The peptide coding domains are located in tandem near the carboxyl terminal end of the precursor and are bounded and separated by paired basic amino acid (lysine-arginine) processing sites.
Function
Neurotensin is involved in a variety of central and peripheral processes. In the brain, it plays a role in modulating hormone activity, social behavior, and learning. For example, neurotensin-expressing neurons in the medial preoptic area (mPOA) of mice project to the ventral tegmental area (VTA), where they contribute to social reward processing and the encoding of odor cues, suggesting a role in both hormonal signaling and reward circuits.[10]
Neurotensin also appears to influence learning processes. In male zebra finches, expression of neurotensin and its receptor genes varies during song development. Both neurotensin and neurotensin receptor mRNA levels decrease during the transition from the sensory to sensorimotor phases of development, implicating neurotensin in the onset of sensorimotor learning. Later in development, neurotensin and neurotensin receptor 1 (Ntsr1) show complementary expression patterns in song-related brain regions, suggesting dynamic modulation of neural responses.[11]
In peripheral tissues, neurotensin is predominantly expressed in the gastrointestinal tract, where it participates in digestion and local signaling. Its aberrant expression has also been associated with tumorigenesis.
Regulation
Neurotensin gene expression is modulated by hormonal and intracellular signaling pathways. In human SK-N-SH neuroblastoma cell cultures and in mice, estrogen has been shown to enhance neurotensin transcription through the activation of cyclic AMP (cAMP) signaling pathways. Estrogen increases cAMP levels and promotes phosphorylation of cAMP response element-binding protein (CREB), a precursor event to neurotensin gene activation. This effect is absent in knockout mice lacking the RIIβ subunit of protein kinase A, highlighting the importance of the cAMP/PKA signaling axis in neurotensin regulation.[12] In female rats, neurotensin mRNA expression peaks in the mPOA during the proestrus phase of the estrous cycle, suggesting regulation by ovarian hormones.[13] Postpartum hormonal states also influence neurotensin and neurotensin receptor expression. In postpartum female mice, neurotensin levels were increased in the paraventricular nucleus (PVN) of the hypothalamus despite reduced Ntsr1 mRNA. Both neurotensin mRNA and peptide levels were elevated in the mPOA, changes that were absent in virgin controls. These patterns are consistent with a regulatory role in maternal behaviors.[14]
Clinical significance
Aberrant neurotensin signaling has been implicated in several pathological conditions, particularly in cancer. In colorectal cancer cells, expression of neurotensin receptor genes (NTSR1 and NTSR2) is regulated by promoter DNA methylation. Downregulation of NTSR1 through RNA interference or pharmacological antagonism results in reduced cell proliferation and migration, indicating a tumor-promoting role for this pathway.[15] Neurotensin expression has also been observed in leiomyomas, or fibroid tumors, of the uterus. Both neurotensin and NTSR1 levels are elevated in fibroid tissues compared to normal uterine tissue, suggesting a role in the pathophysiology of uterine smooth muscle proliferation.[16]
Neurotensin has been implicated in the modulation of dopamine signaling, and produces a spectrum of pharmacological effects resembling those of antipsychotic drugs, leading to the suggestion that neurotensin may be an endogenous neuroleptic. Neurotensin-deficient mice display defects in responses to several antipsychotic drugs consistent with the idea that neurotensin signaling is a key component underlying at least some antipsychotic drug actions.[18] These mice exhibit modest defects in prepulse inhibition (PPI) of the startle reflex, a model that has been widely used to investigate antipsychotic drug action in animals. Antipsychotic drug administration augments PPI under certain conditions. Comparisons between normal and neurotensin-deficient mice revealed striking differences in the ability of different antipsychotic drugs to augment PPI. While the atypical antipsychotic drug clozapine augmented PPI normally in neurotensin-deficient mice, the conventional antipsychotic haloperidol and the newer atypical antipsychotic quetiapine were ineffective in these mice, in contrast to normal mice where these drugs significantly augmented PPI. These results suggest that certain antipsychotic drugs require neurotensin for at least some of their effects. Neurotensin-deficient mice also display defects in striatal activation following haloperidol, but not clozapine administration in comparison to normal wild type mice, indicating that striatal neurotensin is required for the full spectrum of neuronal responses to a subset of antipsychotic drugs.[19]
↑ Friry C, Feliciangeli S, Richard F, Kitabgi P, Rovere C (February 2002). "Production of recombinant large proneurotensin/neuromedin N-derived peptides and characterization of their binding and biological activity". Biochemical and Biophysical Research Communications. 290 (4): 1161–1168. doi:10.1006/bbrc.2001.6308. PMID11811984.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.