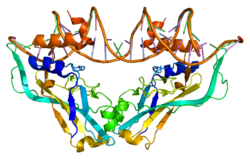
Sonic hedgehog protein(SHH) is encoded for by the SHH gene. The protein is named after the character Sonic the Hedgehog.

Paracrine signaling is a form of cell signaling, a type of cellular communication in which a cell produces a signal to induce changes in nearby cells, altering the behaviour of those cells. Signaling molecules known as paracrine factors diffuse over a relatively short distance, as opposed to cell signaling by endocrine factors, hormones which travel considerably longer distances via the circulatory system; juxtacrine interactions; and autocrine signaling. Cells that produce paracrine factors secrete them into the immediate extracellular environment. Factors then travel to nearby cells in which the gradient of factor received determines the outcome. However, the exact distance that paracrine factors can travel is not certain.
The Wnt signaling pathways are a group of signal transduction pathways which begin with proteins that pass signals into a cell through cell surface receptors. The name Wnt is a portmanteau created from the names Wingless and Int-1. Wnt signaling pathways use either nearby cell-cell communication (paracrine) or same-cell communication (autocrine). They are highly evolutionarily conserved in animals, which means they are similar across animal species from fruit flies to humans.

Zinc finger protein GLI2 also known as GLI family zinc finger 2 is a protein that in humans is encoded by the GLI2 gene. The protein encoded by this gene is a transcription factor.
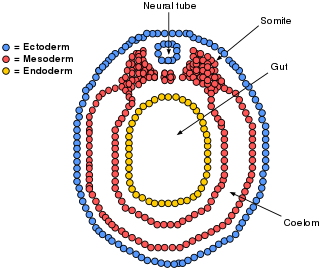
The lateral plate mesoderm is the mesoderm that is found at the periphery of the embryo. It is to the side of the paraxial mesoderm, and further to the axial mesoderm. The lateral plate mesoderm is separated from the paraxial mesoderm by a narrow region of intermediate mesoderm. The mesoderm is the middle layer of the three germ layers, between the outer ectoderm and inner endoderm.
The Hedgehog signaling pathway is a signaling pathway that transmits information to embryonic cells required for proper cell differentiation. Different parts of the embryo have different concentrations of hedgehog signaling proteins. The pathway also has roles in the adult. Diseases associated with the malfunction of this pathway include cancer.
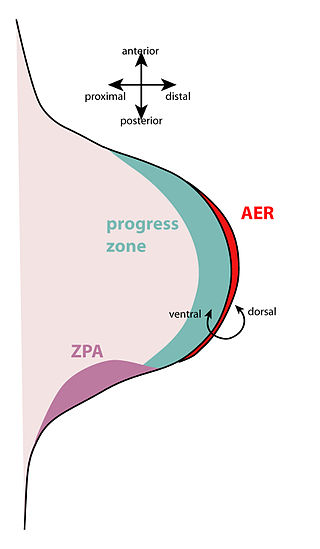
The apical ectodermal ridge (AER) is a structure that forms from the ectodermal cells at the distal end of each limb bud and acts as a major signaling center to ensure proper development of a limb. After the limb bud induces AER formation, the AER and limb mesenchyme—including the zone of polarizing activity (ZPA)—continue to communicate with each other to direct further limb development.

Limb development in vertebrates is an area of active research in both developmental and evolutionary biology, with much of the latter work focused on the transition from fin to limb.
The limb bud is a structure formed early in vertebrate limb development. As a result of interactions between the ectoderm and underlying mesoderm, formation occurs roughly around the fourth week of development. In the development of the human embryo the upper limb bud appears in the third week and the lower limb bud appears four days later.
Gremlin is an inhibitor in the TGF beta signaling pathway. It primarily inhibits bone morphogenesis and is implicated in disorders of increased bone formation and several cancers.
T-box transcription factor TBX5, is a protein that in humans is encoded by the TBX5 gene. Abnormalities in the TBX5 gene can result in altered limb development, Holt-Oram syndrome, Tetra-amelia syndrome, and cardiac and skeletal problems.

Fibroblast growth factor 8(FGF-8) is a protein that in humans is encoded by the FGF8 gene.

Fibroblast growth factor 4 is a protein that in humans is encoded by the FGF4 gene.

T-box transcription factor TBX3 is a protein that in humans is encoded by the TBX3 gene.
The LBH gene is a highly conserved human gene that produces the LBH protein, a transcription co-factor in the Wnt/β-catenin pathway. Upon transcriptional activation of β-catenin, LBH goes on to act as a regulator of cell proliferation and differentiation through multiple transcriptional targets. The gene is located on the p arm of chromosome 2 and is roughly 28 kb long. Current ongoing studies are examining its role in developmental and oncological settings.
Protein odd-skipped-related 1 is a transcription factor that in humans is encoded by the OSR1 gene. The OSR1 and OSR2 transcription factors participate in the normal development of body parts such as the kidney.

Sulfatase 1, also known as SULF1, is an enzyme which in humans is encoded by the SULF1 gene.
The zone of polarizing activity (ZPA) is an area of mesenchyme that contains signals which instruct the developing limb bud to form along the anterior/posterior axis. Limb bud is undifferentiated mesenchyme enclosed by an ectoderm covering. Eventually, the limb bud develops into bones, tendons, muscles and joints. Limb bud development relies not only on the ZPA, but also many different genes, signals, and a unique region of ectoderm called the apical ectodermal ridge (AER). Research by Saunders and Gasseling in 1948 identified the AER and its subsequent involvement in proximal distal outgrowth. Twenty years later, the same group did transplantation studies in chick limb bud and identified the ZPA. It wasn't until 1993 that Todt and Fallon showed that the AER and ZPA are dependent on each other.
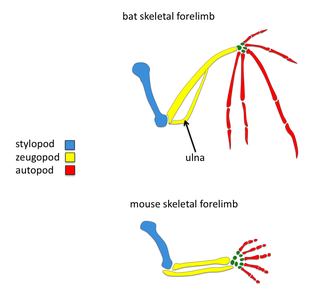
The order Chiroptera, comprising all bats, has evolved the unique mammalian adaptation of flight. Bat wings are modified tetrapod forelimbs. Because bats are mammals, the skeletal structures in their wings are morphologically homologous to the skeletal components found in other tetrapod forelimbs. Through adaptive evolution these structures in bats have undergone many morphological changes, such as webbed digits, elongation of the forelimb, and reduction in bone thickness. Recently, there have been comparative studies of mouse and bat forelimb development to understand the genetic basis of morphological evolution. Consequently, the bat wing is a valuable evo-devo model for studying the evolution of vertebrate limb diversity.
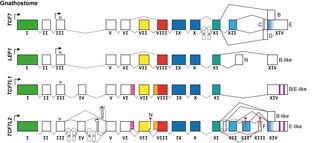
The TCF/LEF family is a group of genes that encode transcription factors which bind to DNA through a SOX-like high mobility group domain. They are involved in the Wnt signaling pathway, particularly during embryonic and stem-cell development, but also had been found to play a role in cancer and diabetes. TCF/LEF factors recruit the coactivator beta-catenin to enhancer elements of genes they target. They can also recruit members of the Groucho family of corepressors.