Antarctic Circumpolar Current (ACC) is an ocean current that flows clockwise from west to east around Antarctica. An alternative name for the ACC is the West Wind Drift. The ACC is the dominant circulation feature of the Southern Ocean and has a mean transport estimated at 100–150 Sverdrups, or possibly even higher, making it the largest ocean current. The current is circumpolar due to the lack of any landmass connecting with Antarctica and this keeps warm ocean waters away from Antarctica, enabling that continent to maintain its huge ice sheet.

Plankton are the diverse collection of organisms that drift in water but are unable to actively propel themselves against currents. The individual organisms constituting plankton are called plankters. In the ocean, they provide a crucial source of food to many small and large aquatic organisms, such as bivalves, fish, and baleen whales.

Coccolithophores, or coccolithophorids, are single-celled organisms which are part of the phytoplankton, the autotrophic (self-feeding) component of the plankton community. They form a group of about 200 species, and belong either to the kingdom Protista, according to Robert Whittaker's five-kingdom system, or clade Hacrobia, according to a newer biological classification system. Within the Hacrobia, the coccolithophores are in the phylum or division Haptophyta, class Prymnesiophyceae. Coccolithophores are almost exclusively marine, are photosynthetic, and exist in large numbers throughout the sunlight zone of the ocean.
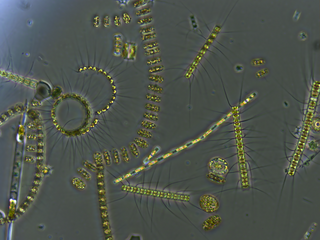
Phytoplankton are the autotrophic (self-feeding) components of the plankton community and a key part of ocean and freshwater ecosystems. The name comes from the Greek words φυτόν, meaning 'plant', and πλαγκτός, meaning 'wanderer' or 'drifter'.

Downwelling is the downward movement of a fluid parcel and its properties within a larger fluid. It is closely related to upwelling, the upward movement of fluid.

Upwelling is an oceanographic phenomenon that involves wind-driven motion of dense, cooler, and usually nutrient-rich water from deep water towards the ocean surface. It replaces the warmer and usually nutrient-depleted surface water. The nutrient-rich upwelled water stimulates the growth and reproduction of primary producers such as phytoplankton. The biomass of phytoplankton and the presence of cool water in those regions allow upwelling zones to be identified by cool sea surface temperatures (SST) and high concentrations of chlorophyll a.
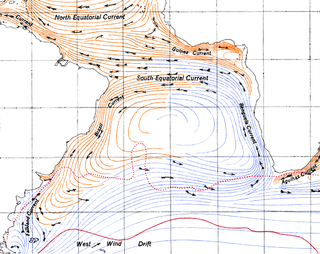
The Benguela Current is the broad, northward flowing ocean current that forms the eastern portion of the South Atlantic Ocean gyre. The current extends from roughly Cape Point in the south, to the position of the Angola-Benguela front in the north, at around 16°S. The current is driven by the prevailing south easterly trade winds. Inshore of the Benguela Current proper, the south easterly winds drive coastal upwelling, forming the Benguela Upwelling System. The cold, nutrient rich waters that upwell from around 200–300 m (656–984 ft) depth in turn fuel high rates of phytoplankton growth, and sustain the productive Benguela ecosystem.
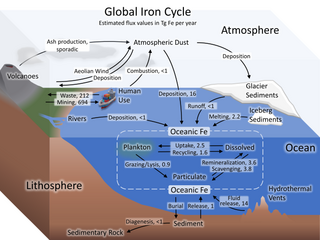
The iron cycle (Fe) is the biogeochemical cycle of iron through the atmosphere, hydrosphere, biosphere and lithosphere. While Fe is highly abundant in the Earth's crust, it is less common in oxygenated surface waters. Iron is a key micronutrient in primary productivity, and a limiting nutrient in the Southern ocean, eastern equatorial Pacific, and the subarctic Pacific referred to as High-Nutrient, Low-Chlorophyll (HNLC) regions of the ocean.

Biogenic silica (bSi), also referred to as opal, biogenic opal, or amorphous opaline silica, forms one of the most widespread biogenic minerals. For example, microscopic particles of silica called phytoliths can be found in grasses and other plants.

Iron fertilization is the intentional introduction of iron-containing compounds to iron-poor areas of the ocean surface to stimulate phytoplankton production. This is intended to enhance biological productivity and/or accelerate carbon dioxide sequestration from the atmosphere. Iron is a trace element necessary for photosynthesis in plants. It is highly insoluble in sea water and in a variety of locations is the limiting nutrient for phytoplankton growth. Large algal blooms can be created by supplying iron to iron-deficient ocean waters. These blooms can nourish other organisms.

Ocean fertilization or ocean nourishment is a type of technology for carbon dioxide removal from the ocean based on the purposeful introduction of plant nutrients to the upper ocean to increase marine food production and to remove carbon dioxide from the atmosphere. Ocean nutrient fertilization, for example iron fertilization, could stimulate photosynthesis in phytoplankton. The phytoplankton would convert the ocean's dissolved carbon dioxide into carbohydrate, some of which would sink into the deeper ocean before oxidizing. More than a dozen open-sea experiments confirmed that adding iron to the ocean increases photosynthesis in phytoplankton by up to 30 times.
John Holland Martin, was an American oceanographer, known for his research work at the Moss Landing Marine Laboratories.

Siliceous ooze is a type of biogenic pelagic sediment located on the deep ocean floor. Siliceous oozes are the least common of the deep sea sediments, and make up approximately 15% of the ocean floor. Oozes are defined as sediments which contain at least 30% skeletal remains of pelagic microorganisms. Siliceous oozes are largely composed of the silica based skeletons of microscopic marine organisms such as diatoms and radiolarians. Other components of siliceous oozes near continental margins may include terrestrially derived silica particles and sponge spicules. Siliceous oozes are composed of skeletons made from opal silica SiO2·nH2O, as opposed to calcareous oozes, which are made from skeletons of calcium carbonate (CaCO3·nH2O) organisms (i.e. coccolithophores). Silica (Si) is a bioessential element and is efficiently recycled in the marine environment through the silica cycle. Distance from land masses, water depth and ocean fertility are all factors that affect the opal silica content in seawater and the presence of siliceous oozes.

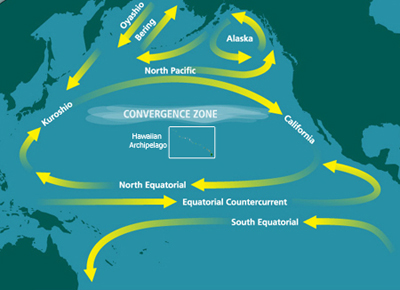
The North Pacific Subtropical Gyre (NPSG) is the largest contiguous ecosystem on earth. In oceanography, a subtropical gyre is a ring-like system of ocean currents rotating clockwise in the Northern Hemisphere and counterclockwise in the Southern Hemisphere caused by the Coriolis Effect. They generally form in large open ocean areas that lie between land masses.

Haida Eddies are episodic, clockwise rotating ocean eddies that form during the winter off the west coast of British Columbia's Haida Gwaii and Alaska's Alexander Archipelago. These eddies are notable for their large size, persistence, and frequent recurrence. Rivers flowing off the North American continent supply the continental shelf in the Hecate Strait with warmer, fresher, and nutrient-enriched water. Haida eddies are formed every winter when this rapid outflow of water through the strait wraps around Cape St. James at the southern tip of Haida Gwaii, and meets with the cooler waters of the Alaska Current. This forms a series of plumes which can merge into large eddies that are shed into the northeast Pacific Ocean by late winter, and may persist for up to two years.
Nutrient cycling in the Columbia River Basin involves the transport of nutrients through the system, as well as transformations from among dissolved, solid, and gaseous phases, depending on the element. The elements that constitute important nutrient cycles include macronutrients such as nitrogen, silicate, phosphorus, and micronutrients, which are found in trace amounts, such as iron. Their cycling within a system is controlled by many biological, chemical, and physical processes.

The viral shunt is a mechanism that prevents marine microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms. The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.
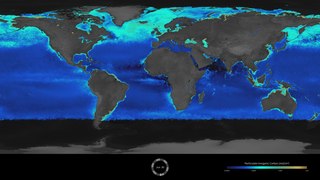
The Great Calcite Belt (GCB) refers to a region of the ocean where there are high concentrations of calcite, a mineral form of calcium carbonate. The belt extends over a large area of the Southern Ocean surrounding Antarctica. The calcite in the Great Calcite Belt is formed by tiny marine organisms called coccolithophores, which build their shells out of calcium carbonate. When these organisms die, their shells sink to the bottom of the ocean, and over time, they accumulate to form a thick layer of calcite sediment.
Low-nutrient, low-chlorophyll (LNLC)regions are aquatic zones that are low in nutrients and consequently have low rate of primary production, as indicated by low chlorophyll concentrations. These regions can be described as oligotrophic, and about 75% of the world's oceans encompass LNLC regions. A majority of LNLC regions are associated with subtropical gyres but are also present in areas of the Mediterranean Sea, and some inland lakes. Physical processes limit nutrient availability in LNLC regions, which favors nutrient recycling in the photic zone and selects for smaller phytoplankton species. LNLC regions are generally not found near coasts, owing to the fact that coastal areas receive more nutrients from terrestrial sources and upwelling. In marine systems, seasonal and decadal variability of primary productivity in LNLC regions is driven in part by large-scale climatic regimes leading to important effects on the global carbon cycle and the oceanic carbon cycle.
Eddy pumping is a component of mesoscale eddy-induced vertical motion in the ocean. It is a physical mechanism through which vertical motion is created from variations in an eddy's rotational strength. Cyclonic (Anticyclonic) eddies lead primarily to upwelling (downwelling). It is a key mechanism driving biological and biogeochemical processes in the ocean such as algal blooms and the carbon cycle.