
Synapsids are one of the two major clades of vertebrate animals in the group Amniota, the other being the sauropsids. The synapsids were the dominant land animals in the late Paleozoic and early Mesozoic, but the only group that survived into the Cenozoic are mammals. Unlike other amniotes, synapsids have a single temporal fenestra, an opening low in the skull roof behind each eye orbit, leaving a bony arch beneath each; this accounts for their name. The distinctive temporal fenestra developed about 318 million years ago during the Late Carboniferous period, when synapsids and sauropsids diverged, but was subsequently merged with the orbit in early mammals.

A therapsid is a member of the clade Therapsida which is a major group of eupelycosaurian synapsids that includes mammals and their ancestors and relatives. Many of the traits today seen as unique to mammals had their origin within early therapsids, including limbs that were oriented more underneath the body, as opposed to the sprawling posture of many reptiles and salamanders.

Dicynodontia is an extinct clade of anomodonts, an extinct type of non-mammalian therapsid. Dicynodonts were herbivores that typically bore a pair of tusks, hence their name, which means 'two dog tooth'. Members of the group possessed a horny, typically toothless beak, unique amongst all synapsids. Dicynodonts first appeared in Southern Pangaea during the mid-Permian, ca. 270–260 million years ago, and became globally distributed and the dominant herbivorous animals in the Late Permian, ca. 260–252 Mya. They were devastated by the end-Permian Extinction that wiped out most other therapsids ca. 252 Mya. They rebounded during the Triassic but died out towards the end of that period. They were the most successful and diverse of the non-mammalian therapsids, with over 70 genera known, varying from rat-sized burrowers to elephant-sized browsers.

Gorgonopsia is an extinct clade of sabre-toothed therapsids from the Middle to Upper Permian roughly 265 to 252 million years ago. They are characterised by a long and narrow skull, as well as elongated upper and sometimes lower canine teeth and incisors which were likely used as slashing and stabbing weapons. Postcanine teeth are generally reduced or absent. For hunting large prey, they possibly used a bite-and-retreat tactic, ambushing and taking a debilitating bite out of the target, and following it at a safe distance before its injuries exhausted it, whereupon the gorgonopsian would grapple the animal and deliver a killing bite. They would have had an exorbitant gape, possibly in excess of 90°, without having to unhinge the jaw.

Suminia is an extinct genus of basal anomodont that lived during the Tatarian age of the late Permian, spanning approximately from 268-252 Ma. Suminia is recognized the youngest non-dicynodont anomodont. Its fossil localities are primarily derived from the Kotel’nich locality of the Kirov Oblast in Russia. However, there have been some isolated specimen found in a few different localities, all from eastern European regions of Russia.
Eothyris is a genus of extinct synapsid in the family Eothyrididae from the early Permian. It was a carnivorous insectivorous animal, closely related to Oedaleops. Only the skull of Eothyris, first described in 1937, is known. It had a 6-centimetre-long (2.4-inch) skull, and its total estimated length was 30 centimetres. Eothyris is one of the most primitive synapsids known and is probably very similar to the common ancestor of all synapsids in many respects. The only known specimen of Eothyris was collected from the Artinskian-lower.

Cotylorhynchus is an extinct genus of herbivorous caseid synapsids that lived during the late Lower Permian (Kungurian) and possibly the early Middle Permian (Roadian) in what is now Texas and Oklahoma in the United States. The large number of specimens found make it the best-known caseid. Like all large herbivorous caseids, Cotylorhynchus had a short snout sloping forward and very large external nares. The head was very small compared to the size of the body. The latter was massive, barrel-shaped, and ended with a long tail. The limbs were short and robust. The hands and feet had short, broad fingers with powerful claws. The barrel-shaped body must have housed large intestines, suggesting that the animal had to feed on a large quantity of plants of low nutritional value. Caseids are generally considered to be terrestrial, though a semi-aquatic lifestyle has been proposed by some authors. The genus Cotylorhynchus is represented by three species, the largest of which could reach more than 6 m in length. However, a study published in 2022 suggests that the genus may be paraphyletic, with two of the three species possibly belonging to separate genera.

Eodicynodon is an extinct genus of dicynodont therapsids, a highly diverse group of herbivorous synapsids that were widespread during the middle-late Permian and early Triassic. As its name suggests, Eodicynodon is the oldest and most primitive dicynodont yet identified, ranging from the middle to late Permian and possessing a mix of ancestral Anomodont/therapsid features and derived dicynodont synapomorphies.

Venyukovia is an extinct genus of venyukovioid therapsid, a basal anomodont from the Middle Permian of Russia. The type and sole species, V. prima, is known only by a partial lower jaw with teeth. Venyukovia has often been incorrectly spelt as 'Venjukovia' in English literature. This stems from a spelling error made by Russian palaeontologist Ivan Efremov in 1940, who mistakenly replaced the 'y' with a 'j', which subsequently permeated through therapsid literature before the mistake was caught and corrected. Venyukovia is the namesake for the Venyukovioidea, a group of small Russian basal anomodonts also including the closely related Otsheria, Suminia, Parasuminia and Ulemica, although it itself is also one of the poorest known. Like other venyukovioids, it had large projecting incisor-like teeth at the front and lacked canines, although the remaining teeth are simple compared to some other venyukovioids, but may resemble those of Otsheria.

Myosaurus is a genus of Anomodontia in the order Therapsida. They are also classified as Dicynodontia, which is a subclade of Anomodontia. The Mysosaurus was a small, herbivorous synapsid that existed around the early Triassic period. All of the fossils found of this species were found in Antarctica and South Africa. Compared to other fossils found from species that existed during this time, the Myosaurus is not common in the fossil record. This is due to a shortage of discovered fossils that possess characteristics unique to the Myosaurus. Notably, under 130 fossil fragments have been found that have been classified as Myosauridae, and almost all have been skulls. These skulls can be classified as Myosaurus because this species, unlike other dicynodonts, do not possess tusks or postfrontal teeth. The only species identified in the family Myosauridae is the Myosaurus gracilis, or M. gracilis. It should be recognized that the Myosaurus is almost always referred to as the M. gracilis in scientific research.

Biseridens is an extinct genus of anomodont therapsid, and one of the most basal anomodont genera known. Originally known from a partial skull misidentified as an eotitanosuchian in 1997, another well-preserved skull was found in the Qingtoushan Formation in the Qilian Mountains of Gansu, China, in 2009 that clarified its relationships to anomodonts, such as the dicynodonts.
Eosyodon is a dubious genus of extinct non-mammalian synapsids from the Permian of Texas. Its type and only species is Eosyodon hudsoni. Though it was originally interpreted as an early therapsid, it is probably a member of Sphenacodontidae, the family of synapsids that includes Dimetrodon.
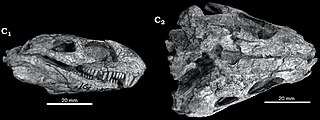
Eosimops is an extinct genus of pylaecephalid dicynodonts. They were small synapsids superficially resembling modern mammals. Eosimops is known from several skull specimens, as well as one complete skeleton. Eosimops lived during the Middle Permian of South Africa.
Knoxosaurus is an extinct genus of non-mammalian synapsids containing the species Knoxosaurus niteckii that existed approximately 279.5 to 268 million years ago. It was named by American paleontologist Everett C. Olson in 1962 on the basis of fragmentary fossils from Middle Permian-age deposits in the San Angelo Formation of Texas in the United States. Olson placed Knoxosaurus in a new infraorder called Eotheriodontia, which he considered a transitional group between the more reptile-like "pelycosaurs" and the more mammal-like therapsids. Knoxosaurus and Olson's other eotheriodonts were later considered to be undiagnostic remains of basal synapsids, no more closely related to therapsids than are other pelycosaur-grade synapsids.

Aulacephalodon is an extinct genus of medium-sized dicynodonts, or non-mammalian synapsids, that lived during late Permian period. Individuals of Aulacephalodon are commonly found in the Lower Beaufort Group of the Karoo Supergroup of South Africa. Rising to dominance during the Late Permian, Aulacephalodon was among the largest terrestrial vertebrate herbivores until its extinction at the end of the Permian. Two species have been named, the type species, A. bainii, and a second species, A. peavoti. However, debate exists among paleontologists if A. peavoti is a true member of the genus Aulacephalodon. Aulacephalodon belongs to the family Geikiidae, a family of dicynodonts generally characterized by their short, broad skulls and large nasal bosses. Sexual dimorphism has been identified in A. bainii.

Chainosauria is a large and speciose clade of anomodont therapsid that includes the highly diverse dicynodonts and a small number of closely related basal genera —although the total composition and taxonomic scope of Chainosauria is in flux. Chainosauria was named in 1923 to group together the dicynodonts and their close relatives, namely three small anomodont genera from South Africa that made up the now defunct group 'Dromasauria'. The name soon fell into disuse, however, as it was functionally replaced by Anomodontia. Chainosauria was later revived cladistically in 2009, preserving the association of dicynodonts and the 'dromasaurs' and has since served in effect as both a cladistic and a biogeographic counterpart to the Laurasian venyukovioids, with early chainosaurs appearing to have been a Gondwanan radiation.
Kunpania is an extinct genus of dicynodont therapsid from the Quanzijie Formation of Xinjiang, China. The type and only species is K. scopulusa, and it is known only by a single incomplete specimen including parts of the skull and forelimb. Since its initial description in 1978 by palaeontologist Ailing Sun, it has sometimes been considered to be another species of Dicynodon by other researchers, or potentially undiagnostic. However, a redescription in 2021 reaffirmed its distinctiveness, including a uniquely well developed muscle attachment on the humerus. Kunpania is perhaps the oldest known member of the derived dicynodont group Dicynodontoidea, potentially dating to the Middle Permian period during the Capitanian, and so may fill a knowledge gap in the history of dicynodont evolution.
Gorgodon is an extinct genus of basal synapsids. The genus is monotypic, known only from the type species Gorgodon minutus from the Early Permian of the southwestern United States. The only known remains of Gorgodon are two fossils consisting of fragments of the skull. Gorgodon was described and named by paleontologist Everett C. Olson in 1962 from the San Angelo Formation in Knox County, Texas. Based on what is known of Gorgodon—the squamosal, quadrate, and pterygoid bones of the back of the skull, the maxilla and premaxilla bones that make up the front of the skull, and several teeth—Gorgodon had a relatively large temporal fenestra and a pair large, conical caniniform teeth at the front of the jaw. Other distinguishing features of Gorgodon include the fused connection between the quadrate and squamosal bones and a long transverse process of the pterygoid.

Anomocephaloidea is a clade of basal anomodont therapsids related to the dicynodonts known from what is now South Africa and Brazil during the Middle Permian. It includes only two species, Anomocephalus africanus from the Karoo Basin of South Africa and Tiarajudens eccentricus from the Paraná Basin of Brazil. Anomocephaloidea was named in 2011 with the discovery of Tiarajudens, although Anomocephalus itself has been known since 1999.
Taoheodon is an extinct genus of dicynodont therapsid from the Sunjiagou Formation in the Shanxi province of China, dated to the Wuchiapingian age of the Late Permian. Its type and only known species is T. baizhijuni. Taoheodon was a close relative of the well known Dicynodon, and may represent a biogeographical link between the South African Dicynodon and similar dicynodonts found in Laos.






