Petrolacosaurus is an extinct genus of diapsid reptile from the late Carboniferous period. It was a small, 40-centimetre (16 in) long reptile, and one of the earliest known reptile with two temporal fenestrae. This means that it was at the base of Diapsida, the largest and most successful radiation of reptiles that would eventually include all modern reptile groups, as well as dinosaurs and other famous extinct reptiles such as plesiosaurs, ichthyosaurs, and pterosaurs. However, Petrolacosaurus itself was part of Araeoscelida, a short-lived early branch of the diapsid family tree which went extinct in the mid-Permian.

Caseasauria is one of the two main clades of early synapsids, the other being the Eupelycosauria. Caseasaurs are currently known only from the Late Carboniferous and the Permian, and include two superficially different families, the small insectivorous or carnivorous Eothyrididae, and the large, herbivorous Caseidae. These two groups share a number of specialised features associated with the morphology of the snout and external naris.
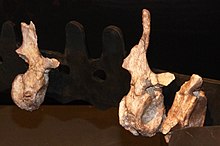
Caseidae are an extinct family of basal synapsids that lived from the Late Carboniferous to Middle Permian between about 300 and 265 million years ago. Fossils of these animals come from the south-central part of the United States, from various parts of Europe, and possibly from South Africa if the genus Eunotosaurus is indeed a caseid as some authors proposed in 2021. Caseids show great taxonomic and morphological diversity. The most basal taxa were small insectivorous and omnivorous forms that lived mainly in the Upper Carboniferous and Lower Permian, such as Eocasea, Callibrachion, and Martensius. This type of caseid persists until the middle Permian with Phreatophasma and may be Eunotosaurus. During the early Permian, the clade is mainly represented by many species that adopted a herbivorous diet. Some have evolved into gigantic forms that can reach 6–7 metres (20–23 ft) in length, such as Cotylorhynchus hancocki and Alierasaurus ronchii, making them the largest Permian synapsids. Caseids are considered important components of early terrestrial ecosystems in vertebrate history because the numerous herbivorous species in this family are among the first terrestrial tetrapods to occupy the role of primary consumer. The caseids experienced a significant evolutionary radiation at the end of the early Permian, becoming, with the captorhinid eureptiles, the dominant herbivores of terrestrial ecosystems in place of the edaphosaurids and diadectids.
Casea is a genus of herbivorous caseid synapsids that lived during the late Lower Permian (Kungurian) in what is now Texas, United States. The genus is only represented by its type species, Casea broilii, named by Samuel Wendell Williston in 1910. The species is represented by a skull associated with a skeleton, a second skull, a partial skull with a better preserved dentition than that of the preceding skulls, and several incomplete postcranial skeletons. Three other Casea species were later erected, but these are considered today to be invalid or belonging to different genera. Casea was a small animal with a length of about 1.20 m and a weight of around 20 kg.

Oedaleops is an extinct genus of caseasaur synapsids from the Early Permian of the southwestern United States. Fossils have been found in the Cutler Formation in New Mexico, which dates back to the Wolfcampian stage of the Early Permian. All remains belong to the single known species Oedaleops campi. Oedaleops was closely related to Eothyris, and both are part of the family Eothyrididae. Like Eothyris, it was probably an insectivore.

Cotylorhynchus is an extinct genus of herbivorous caseid synapsids that lived during the late Lower Permian (Kungurian) and possibly the early Middle Permian (Roadian) in what is now Texas and Oklahoma in the United States. The large number of specimens found make it the best-known caseid. Like all large herbivorous caseids, Cotylorhynchus had a short snout sloping forward and very large external nares. The head was very small compared to the size of the body. The latter was massive, barrel-shaped, and ended with a long tail. The limbs were short and robust. The hands and feet had short, broad fingers with powerful claws. The barrel-shaped body must have housed large intestines, suggesting that the animal had to feed on a large quantity of plants of low nutritional value. Caseids are generally considered to be terrestrial, though a semi-aquatic lifestyle has been proposed by some authors. The genus Cotylorhynchus is represented by three species, the largest of which could reach more than 6 m in length. However, a study published in 2022 suggests that the genus may be paraphyletic, with two of the three species possibly belonging to separate genera.
Angelosaurus is an extinct genus of herbivorous caseid synapsids that lived during the late Lower Permian (Kungurian) and early Middle Permian (Roadian) in what is now Texas and Oklahoma in the United States. Like other herbivorous caseids, it had a small head, large barrel-shaped body, long tail, and massive limbs. Angelosaurus differs from other caseids by the extreme massiveness of its bones, particularly those of the limbs, which show a strong development of ridges, processes, and rugosities for the attachment of muscles and tendons. Relative to its body size, the limbs of Angelosaurus were shorter and wider than those of other caseids. The ungual phalanges looked more like hooves than claws. The few known cranial elements show that the skull was short and more robust than that of the other representatives of the group. Angelosaurus is also distinguished by its bulbous teeth with shorter and wider crowns than those of other caseids. Their morphology and the high rate of wear they exhibit suggests a diet quite different from that of other large herbivorous caseids, and must have been based on particularly tough plants. A study published in 2022 suggests that the genus may be paraphyletic, with Angelosaurus possibly only represented by its type species A. dolani.

Ennatosaurus is an extinct genus of caseid synapsid that lived during the Middle Permian in northern European Russia. The genus is only represented by its type species, Ennatosaurus tecton, which was named in 1956 by Ivan Antonovich Efremov. The species is known from at least six skulls associated with their lower jaws, as well as from the postcranial bones of several juvenile individuals. Ennatosaurus has the typical caseid skull with a short snout tilted forward and very large external nares. However, it differs from other derived caseids by its postcranial skeleton with smaller proportions compared to the size of the skull. As with other advanced caseids, the teeth of Ennatosaurus were well suited for slicing and cutting vegetation. The presence of a highly developed hyoid apparatus indicates the presence of a massive and mobile tongue, which had to work in collaboration with the palatal teeth during swallowing. With a late Roadian - early Wordian age, Ennatosaurus is one of the last known caseids.

Qianosuchus is an extinct genus of aquatic poposauroid archosaur from the middle Triassic (Anisian) Guanling Formation of Pan County, China. It is represented by two nearly complete skeletons and a crushed skull preserved in the limestone. Qianosuchus was at least 3 metres long, and had several skeletal adaptations which indicate a semi-marine lifestyle, similar to modern-day saltwater crocodiles. These adaptations have not been seen in any other archosaur from the Triassic.
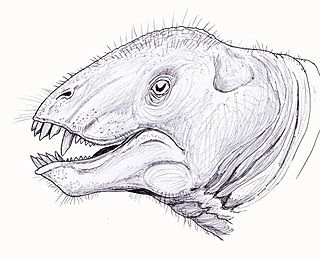
Tapinocaninus is an extinct genus of therapsids in the family Tapinocephalidae, of which it is the most basal member. Only one species is known, Tapinocaninus pamelae. The species is named in honor of Rubidge's mother, Pam. Fossils have been found dating from the Middle Permian.

Tetragonias is an extinct genus of dicynodont from the Anisian Manda Beds of Tanzania. With tetra meaning “four,” and goni meaning “angle,” the name references the square shape of the Tetragonias skull when viewed dorsally. Not to be confused with the plant Tetragonia,Tetragonias were dicynodont anomodonts discovered in the late 1960s by paleontologist A. R. I. Cruickshank in the Manda Formation. Only the type species, T. njalilus, has been recognized.

Heleosaurus scholtzi is an extinct species of basal synapsids, known as pelycosaurs, in the family of Varanopidae during the middle Permian. At first H. scholtzi was mistakenly classified as a diapsid. Members of this family were carnivorous and had dermal armor, and somewhat resembled monitor lizards. This family was the most geologically long lived, widespread, and diverse group of early amniotes. To date only two fossils have been found in the rocks of South Africa. One of these fossils is an aggregation of five individuals.

Callibrachion is an extinct genus of caseid synapsids that lived in east-central France during the Lower Permian (Asselian). The holotype and only known specimen (MNHN.F.AUT490) is represented by an almost complete postcranial skeleton associated with skull fragments discovered at the end of the 19th century in the Permian Autun basin in Saône-et-Loire department, in the Bourgogne-Franche-Comté region. It belongs to an immature individual measuring less than 1.50 m in length. Callibrachion was long considered a junior synonym of the genus Haptodus and classified among the sphenacodontid pelycosaurs. In 2015, a new study found that Callibrachion was a different animal from Haptodus and that it was a caseasaur rather than a sphenacodontid. This was confirmed in 2016 by a cladistic analysis which recovered Callibrachion as a basal caseid. Callibrachion's sharp teeth and unenlarged ribcage indicate that this animal was likely faunivorous.
Ruthenosaurus is an extinct genus of caseid synapsids that lived in what is now southern France during the Early Permian about 285 million years ago. It is known from the holotype MNHN.F.MCL-1 an articulated partial postcranial skeleton. It was collected by D. Sigogneau-Russell and D. Russell in 1970 in the upper part of the M2 Member, Grès Rouge Group, in the Rodez Basin, near the village of Valady, in Occitanie Region. It was first named by Robert R. Reisz, Hillary C. Maddin, Jörg Fröbisch and Jocelyn Falconnet in 2011, and the type species is Ruthenosaurus russellorum.

Euromycter is an extinct genus of caseid synapsids that lived in what is now southern France during the Early Permian about 285 million years ago. The holotype and only known specimen of Euromycter (MNHN.F.MCL-2) includes the complete skull with lower jaws and hyoid apparatus, six cervical vertebrae with proatlas, anterior part of interclavicle, partial right clavicle, right posterior coracoid, distal head of right humerus, left and right radius, left and right ulna, and complete left manus. It was collected by D. Sigogneau-Russell and D. Russell in 1970 at the top of the M1 Member, Grès Rouge Group, near the village of Valady, Rodez Basin. It was first assigned to the species "Casea" rutena by Sigogneau-Russell and Russell in 1974. More recently, it was reassigned to its own genus, Euromycter, by Robert R. Reisz, Hillary C. Maddin, Jörg Fröbisch and Jocelyn Falconnet in 2011. The preserved part of the skeleton suggests a size between 1,70 m (5,5 ft) and 1,80 m (5,9 ft) in length for this individual.

Pampaphoneus is an extinct genus of carnivorous dinocephalian therapsid belonging to the family Anteosauridae. It lived 268 to 265 million years ago during the Wordian age of the Guadalupian period in what is now Brazil. Pampaphoneus is known by an almost complete skull with the lower jaw still articulated, discovered on the lands of the Boqueirão Farm, near the city of São Gabriel, in the state of Rio Grande do Sul. A second specimen from the same locality was reported in 2019 and 2020 but has not yet been described. It is composed of a skull associated with postcranial remains. It is the first South American species of dinocephalian to have been described. The group was previously known in South America only by a few isolated teeth and a jaw fragment reported in 2000 in the same region of Brazil. Phylogenetic analysis conducted by Cisneros and colleagues reveals that Pampaphoneus is closely related to anteosaurs from European Russia, indicating a closer faunal relationship between South America and Eastern Europe than previously thought, thus promoting a Pangea B continental reconstruction.

Ichthyovenator is a genus of spinosaurid dinosaur that lived in what is now Laos, sometime between 125 and 113 million years ago, during the Aptian stage of the Early Cretaceous period. It is known from fossils collected from the Grès supérieurs Formation of the Savannakhet Basin, the first of which were found in 2010, consisting of a partial skeleton without the skull or limbs. This specimen became the holotype of the new genus and species Ichthyovenator laosensis, and was described by palaeontologist Ronan Allain and colleagues in 2012. The generic name, meaning "fish hunter", refers to its assumed piscivorous lifestyle, while the specific name alludes to the country of Laos. In 2014, it was announced that more remains from the dig site had been recovered; these fossils included teeth, more vertebrae (backbones) and a pubic bone from the same individual.

Alierasaurus is an extinct genus of caseid synapsid that lived during the early Middle Permian (Roadian) in what is now Sardinia. It is represented by a single species, the type species Alierasaurus ronchii. Known from a very large partial skeleton found within the Cala del Vino Formation, Alierasaurus is one of the largest known caseids. It closely resembles Cotylorhynchus, another giant caseid from the San Angelo Formation in Texas. The dimensions of the preserved foot elements and caudal vertebrae suggest an estimated total length of about 6 or 7 m for Alierasaurus. In fact, the only anatomical features that differ between Alierasaurus and Cotylorhynchus are found in the bones of the feet; Alierasaurus has a longer and thinner fourth metatarsal and it has ungual bones at the tips of the toes that are pointed and claw-like rather than flattened as in other caseids. Alierasaurus and Cotylorhynchus both have very wide, barrel-shaped rib cages indicating that they were herbivores that fed primarily on high-fiber plant material.

Arisierpeton is an extinct genus of synapsids from the Early Permian Garber Formation of Richards Spur, Oklahoma. It contains a single species, Arisierpeton simplex.

Cabarzia is an extinct genus of varanopid from the Early Permian of Germany. It contains only a single species, Cabarzia trostheidei, which is based on a well-preserved skeleton found in red beds of the Goldlauter Formation. Cabarzia shared many similarities with Mesenosaurus romeri, although it did retain some differences, such as more curved claws, a wide ulnare, and muscle scars on its sacral ribs. With long, slender hindlimbs, a narrow body, an elongated tail, and short, thick forelimbs, Cabarzia was likely capable of running bipedally to escape from predators, a behavior shared by some modern lizards. It is the oldest animal known to have adaptations for bipedal locomotion, predating Eudibamus, a bipedal bolosaurid parareptile from the slightly younger Tambach Formation.