Left: Illustration of N.bicalcarata from Macfarlane's 1908 monograph, showing the hollow tendrils Right: Upper pitcher and part of male inflorescence of N.dyak, from Moore's 1880 description[3]
Nepenthes bicalcarata was introduced to Europe in 1879 by British explorer Frederick William Burbidge, who collected plants for the famous Veitch Nursery. These were cultivated to larger size and distributed in 1881.[4]
During this time, interest in Nepenthes had reached its peak. A note in The Gardeners' Chronicle of 1881 mentions the Veitch Nursery's N.bicalcarata as follows:[12]
"Then there is N.bicalcarata, a most robust habited kind with sturdy foliage and bag-like pitchers provided with a vicious-looking rat-trap-like apparatus in its lid which renders it very distinct from its neighbours."
Several years after its introduction, N.bicalcarata was still very much a horticultural rarity. In Veitch's catalogue for 1889, N.bicalcarata was priced at £3.3s per plant, while the famous giant-pitchered N.northiana and N.rajah were selling for £2.2s.[4]
Description
Nepenthes bicalcarata plants are the largest in the genus, climbing up to 20 m into the forest canopy. The cylindrical stem is thicker than that of any other Nepenthes species, measuring up to 3.5cm in diameter. Internodes are up to 40cm long.[11][13]
Climbing plant with upper pitchers
The leaves of N.bicalcarata are petiolate and coriaceous in texture. The lamina is obovate-lanceolate in form and also reaches huge dimensions, growing to 80cm in length and 12cm in width.[14] It is slightly decurrent on the stem, forming two narrow wings. The lamina has indistinct longitudinal veins and numerous pennate veins. Tendrils may be up to 60cm long and 8mm wide. They are hollow and swollen near the pitcher.[11]
Lower pitchers
Although most parts of the plant are very large, the pitchers themselves do not rival those of species such as N.rajah. Nevertheless, they may have a volume of over one litre and grow up to 25cm high and 16cm wide. A pair of prominent fringed wings (≤15mm wide) runs down the front of lower pitchers. These are usually reduced to ribs in aerial pitchers. The peristome (≤20mm wide) is characteristically flattened and curved inwards.[11] The inner portion of the peristome accounts for around 70% of its total cross-sectional surface length.[15] It bears small but distinct teeth. The two sharp spines for which the species is famous are present on the underside of the pitcher lid, and may be 3cm long. They are derived from the uppermost 10–12 peristome ribs.[16] The pitcher lid or operculum is reniform to cordate and has no appendages. An unbranched spur (≤15mm long) is inserted at the base of the lid.[11]
Nepenthes bicalcarata has a paniculateinflorescence. The peduncle may be up to 40cm long and the rachis can reach 100cm in length. Female inflorescences are usually shorter.[17] Branches on the flower stem are up to 40mm long and bear up to 15 flowers. Sepals are either obovate or lanceolate and up to 4mm long.[11] A study of 120 pollen samples taken from a herbarium specimen (Fosberg 43860, altitude not recorded) found the mean pollen diameter to be 28.9 μm (SE = 0.4; CV = 7.5%).[18]
Mature plants are virtually glabrous. Caducous hairs are present on the youngest parts of the plant and on the inflorescences.
Nepenthes bicalcarata varies little across its range. Consequently, no infraspecific taxa have been described.[11]
Specimens growing in undisturbed peat swamp forest, where sunlight is greatly diffused and high humidity prevails, reach the largest dimensions.[11]Nepenthes bicalcarata has a shallow root system that only penetrates the top layer of peat and leaf litter, to a depth of about 25cm. Below this, high concentrations of tannins and alkaloids render the substrate toxic.[4]
Nepenthes bicalcarata is generally found below 300m in altitude, although Johannes Gottfried Hallier reported a single collection in 1894 from between 700 and 950m above sea level.[19]
The two thorns that give N.bicalcarata its name are unique to this species and bear some of the largest nectaries in the plant kingdom.[21][22][23] The purpose of these structures has long been debated among botanists. Frederick William Burbidge suggested that they might serve to deter arborealmammals such as tarsiers, lorises and monkeys from stealing the contents of the pitchers.[11][24] In an article published in 1982, Cliff Dodd speculated on the function of the thorns, but did not believe they play a role in prey capture.[25]Charles Clarke observed that monkeys and tarsiers rip the pitchers open at the sides in order to feed from them, rather than reaching in through the pitcher mouth.[26] However, he found that the mammals attacked the pitchers of N.bicalcarata less frequently than those of other species, such as N.rafflesiana.[26] Clarke's observations suggest that the spines likely serve to lure insects into a precarious position over the pitcher mouth, where they may lose their footing and fall into the pitcher fluid, eventually drowning.[26] A similar trapping method is employed by the Sumatran endemic N.lingulata, which has a single filiform appendage positioned over the pitcher mouth. However, in that species the appendage is a structure of the lid and not the peristome.[27]
Together with N.ampullaria and N.ventricosa, N.bicalcarata is unusual in that the glandular region of the pitcher extends almost to the peristome, such that there is little or no conductive waxy zone.[15][28][29][30] The waxy zone functions by causing prey to slip and fall into the digestive fluid. A 2004 study found that the peristome of N.bicalcarata plays a very important role in prey capture. When it is dry, the peristome is highly ineffective in catching insect prey, but when wetted, the capture rate increases more than three-fold.[28]
A 1999 study of pitcher morphology and spectral reflectance characteristics showed that N.bicalcarata has no colour contrast maximum between the peristome and pitcher cup in the ultraviolet waveband, but small maxima of –0.17 at 450nm (violet), 0.32 at 548nm (green), and –0.16 at 668nm (red).[31] This means that the peristome is less reflective than the pitcher cup in the violet and red bands, but more reflective in the green band. The contrast maxima in the green and blue regions seem to correspond to insect visual sensitivity maxima, while the others do not.[31] Based on their findings, the authors made the "tentative prediction" that the upper pitchers of N.bicalcarata are less successful at catching anthophilous (flower-visiting) prey than those of sympatric N.rafflesiana.[31]
Association with ants and pitcher infauna
Nepenthes bicalcarata plays host to an unusual species of ant that makes its nest in the plant's hollow tendrils.[11][32][33][34] Described as Camponotus schmitzi in 1933,[35] it is a member of the extremely populous and widespread genus of carpenter ants.
Intermediate pitcher with swollen tendril colonised by Camponotus schmitzi.
This unique animal–plant interaction was noted by Frederick William Burbidge as early as 1880.[36] In 1904, Odoardo Beccari suggested that the ants feed on insects found on and around the plant, but may fall prey to it themselves.[37] In 1990, B. Hölldobler and E. O. Wilson proposed that N.bicalcarata and C.schmitzi form a mutually beneficial association.[38] At the time, however, no experimental data existed to support such a hypothesis. A series of observations and experiments carried out in Brunei by Charles Clarke (published in 1992 and 1998),[39][40][41] and by Clarke and Roger Kitching (1993 and 1995),[42][43] strongly support the mutualism theory.
Nepenthes bicalcarata is a myrmecotroph (ant-fed plant), obtaining nutrients from C.schmitzi in the form of egesta and, occasionally, ant remains. It has been estimated that this input accounts for 42% of the plant's total foliar nitrogen (76% in plants with ant occupancy rates above 75%).[5] The ants increase nutrient retention in the pitchers by preying on infaunal flies, which would otherwise eventually leave their hosts and thereby act as kleptoparasites. This nutrients later becomes available to the plant through the ants' waste.[44]Camponotus schmitzi has also been observed to attack newly caught insects and therefore prevent prey escape.[45][46] At other times, the ants are very passive, remaining hidden under the inner peristome fold, presumably so as not to dissuade visitation by potential prey species.[45] This behaviour is in stark contrast to other myrmecophytic ants, which are typically highly territorial.[45] John Thompson has suggested that N.bicalcarata may be the only plant species that obtains nutrients through both insect capture and ant-hosting habits.[47]
Camponotus schmitzi is able to swim in the pitcher fluid using tripod-like leg coordination similar to that of terrestrial locomotion[48][49] and can remain submerged for up to 30 seconds.[50] When feeding, it appears to target large prey items only, cooperatively retrieving them from the fluid.[50] Hauling food from the pitcher fluid to the peristome—a distance of no more than 5cm—may take up to 12 hours.[11] In this way the contents of N.bicalcarata pitchers is controlled such that organic matter does not accumulate to the point of putrefaction, which could lead to the demise of pitcher infauna (which may also benefit the plant) and sometimes the pitcher itself.[11][51]
The ants have been observed to clean the peristome of fungal hyphae and other contaminants, thereby maintaining high trapping efficiency over the pitcher's lifespan.[52][53] Research conducted by Dennis and Marlis Merbach has shown that C.schmitzi also benefits N.bicalcarata by protecting it from pitcher-destroying weevils of the genus Alcidodes.[54][55] In order to create a favourable environment for its pitcher inhabitants, it appears that N.bicalcarata actively maintains the pH of its pitcher fluid at a less acidic level than that found in most other Nepenthes species[29][45] (this might explain the occasional presence of tree frog eggs in its pitchers).[29] In doing so, however, the plant reduces its ability to digest and assimilate nutrients from captured prey. The pitcher fluid of N.bicalcarata is also less viscoelastic than that of most Nepenthes species, and appears to lack functional digestive enzymes.[5]Nepenthes bicalcarata is therefore highly reliant on its ant symbiont. Indeed, plants not inhabited by C.schmitzi do not appear to benefit significantly from carnivory, with any gains from prey digestion being offset by the high costs of pitcher construction.[5] Conversely, ant-inhabited plants have more leaves and a greater total leaf area, and ant presence is associated with lower pitcher abortion rates and more voluminous pitchers (and consequently greater prey biomass).[5][56][57] Studies have shown that plants colonised by C.schmitzi have more nitrogen available to them, and a higher proportion of it is insect-derived.[44]
Camponotus schmitzi nests solely in the tendrils of N.bicalcarata and rarely ventures onto other plants. The species is completely dependent on N.bicalcarata for food and domicile.[11]Nepenthes bicalcarata, on the other hand, is able to survive and reproduce without the presence of the ants; it is a facultative mutualist. This being the case, there appear to be few mature plants over 2 metres in height not colonised by C.schmitzi.[11] The ants seem to favour upper pitchers and rarely colonise lower pitchers.[58] This is likely due to the fact that terrestrial traps are periodically submerged in water during heavy rains. Flooding of the ants' nest chamber could result in the demise of the ant colony, particularly the developing eggs, larvae, and pupae.[23]
A species of mite, Naiadacarus nepenthicola, appears to be restricted to the pitchers of N.bicalcarata. It is thought to feed on decomposing leaves and insects that are caught in the pitchers. Deutonymphs of this mite are dispersed through phoresy on C.schmitzi.[59]
Summary of results from study by Bazile et al. (2012)[5]
Left: Relationship between total foliar area and plant height in ant-inhabited specimens (PA), pitchering specimens lacking ants (PnoA), and non-pitchering specimens (NoP). The highlighted value of 175cm indicates the approximate height at which plants transition from a self-supporting stem with lower pitchers (filled points) to a climbing one with upper pitchers (empty points).[5]
Left centre: Effect of C.schmitzi occupancy on leaf apex abortion and pitcher production rates. In the first chart (A), cases where the tendril was found to be cut are grouped under unknown fate (denoted with a question mark) and "pitcher" encompasses both living and dead traps (in non-pitchering plants, the latter).[5] Right centre: Prey biomass accumulated over a pitcher's entire lifespan as a function of pitcher volume in ant-occupied and unoccupied lower pitchers.[5]
Right:A: Isotopic signatures (δ15N) of C.schmitzi, ant-occupied plants (PA), and unoccupied plants with no evidence of previous colonisation (PnoA-no hole). B: Relationship between C.schmitzi occupation rate and plant foliar δ15N.[5]
Summary of results from study by Scharmann et al. (2013)[44]
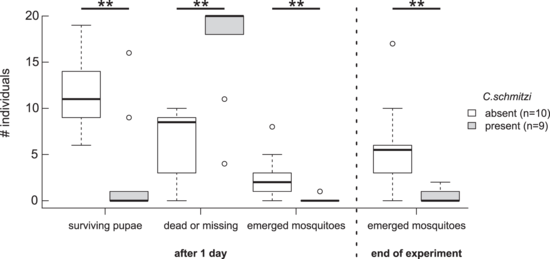
Left: Natural nitrogen isotope abundance ratio (δ15N) of ant-occupied and unoccupied N.bicalcarata plants (highlighted in grey), associated food web components, and non-carnivorous plants ("non-CPs"). The dotted horizontal line indicates the median δ15N of prey insects.[44] Centre: Nitrogen flux from C.schmitzi colonies to an N.bicalcarata host plant (c. 1.4m diameter). The bars show the change in 15N concentration in the host plant's leaves two weeks after a pulse of 15N was fed to the associated C.schmitzi colonies. Leaf node 1 has the youngest (still developing) leaf, with the others numbered sequentially as one moves down the stem. The pictograms below the graph show the state of each leaf and pitcher, and indicate the presence or absence of C.schmitzi and whether a 15N pulse was administered.[44] Right: Effect of C.schmitzi presence on survival of infaunal mosquito pupae and successful emergence of mosquitoes (Aedes sp.) from N.bicalcarata pitchers. The experiment began with 20 living pupae occupying each pitcher.[44]
Camponotus schmitzi ants hunting mosquito pupae in a pitcher of N.bicalcarata
Camponotus schmitzi ant hunting a large fly larva of the genus Eristalis in a pitcher of N.bicalcarata
Natural hybrids
The following natural hybrids involving N.bicalcarata have been recorded.
None of these hybrids are known to be colonised by C. schmitzi as is the parent species N.bicalcarata.[11]
N.ampullaria × N.bicalcarata
Lower pitcher of N.ampullaria × N.bicalcarata (cultivated)
Since N.ampullaria and N.bicalcarata are often sympatric in the wild, it is not surprising that they occasionally hybridise. Several examples of this natural hybrid are known from Brunei, where it grows in peat swamp forest and heath forest.[11] It was first recorded by Mitsuru Hotta in 1966.[61]
Pitchers bear the characteristic thorns of N.bicalcarata, although they are greatly reduced in size. The mouth is round and the lid is ovate-cordate in form. This hybrid resembles N. × hookeriana to a certain extent, but differs in having spines on the underside of the lid and more globose upper pitchers with a smaller lid.[11]
Aerial pitchers of N.ampullaria × N.bicalcarata are usually dominated by characteristics of N.ampullaria. They are often very small and unable to function in a normal manner.[11]
(N.ampullaria × N.gracilis) × N.bicalcarata
This rare complex hybrid was discovered in the lowland forests of Brunei. It produces small speckled pitchers with reduced thorns under the lid. The plant is intermediate in appearance between N. × trichocarpa and N.bicalcarata.
This hybrid has been named N.×cantleyi after Rob Cantley. The morphology of the pitchers closely resembles N.bicalcarata, although the peristome is more similar to that of N.gracilis. The characteristic spines of N.bicalcarata are greatly reduced and are only present as small bumps. This hybrid has the growth habit of N.gracilis, with the stem scrambling along the ground. It grows in open, sandy areas. The pitcher fluid is notably acidic like that of N.gracilis, with pH values as low as 1.82 being recorded.[11]
N.bicalcarata × N.mirabilis var. echinostoma
This natural hybrid is rare and is only known from Brunei, where it involves N.mirabilis var. echinostoma. It closely resembles N.bicalcarata, although it differs from that species in having reduced thorns and a wider peristome. Unlike N.bicalcarata, this hybrid inhabits open areas with sandy substrates.[11]
N.bicalcarata × N.rafflesiana
This relatively rare natural hybrid is sometimes found growing among populations of N.bicalcarata and N.rafflesiana. It is intermediate in form between its parent species.
(N.bicalcarata × N.rafflesiana) × N.mirabilis var. echinostoma
Charles Clarke reported a single plant displaying characters intermediate between those of N.bicalcarata, N.rafflesiana and N.mirabilis var. echinostoma, and growing among populations of these species. It is likely a complex hybrid, although this cannot be proven without the use of molecular techniques.[11]
Cultivation and cultivars
Lower pitcher of a plant cultivated in Kuching, Sarawak.
Nepenthes bicalcarata may be cultivated in a similar manner to other members of the genus, though it requires relatively hot and humid conditions.[25][62] It needs large pots to do well, and temporary halts in growth are often due to becoming root-bound. Re-potting to a larger pot will often result in a rapid size increase. Under satisfactory conditions, this plant will grow quite fast for a Nepenthes, reaching a large size relatively rapidly.
Despite naturally occurring in peat swamp forests, N.bicalcarata has been successfully grown in a completely inorganic substrate consisting of one part each of Seramis clay perls, lava gravel, and Lecaton expanded clay perls.[63]
Nepenthes bicalcarata has been artificially crossed with N.hirsuta. A particularly robust-pitchered cultivar of this hybrid was named Nepenthes 'Hortulanus Otten', in honour of Karel Otten, former curator of the Botanic Garden in Ghent, Belgium.[64]
A cultivar of the artificial cross N.bicalcarata × N. × dyeriana was registered in 1988 under the name Nepenthes 'Nina Dodd'. It is named after a relative of Cliff Dodd, who created the hybrid.[65]
Two further cultivars of manmade hybrids involving N.bicalcarata have been named. These are Nepenthes 'Bella' ((N.ampullaria × N.bicalcarata) × N.truncata) and Nepenthes 'Rapa' (N.ampullaria × N.bicalcarata).[66] However, both of these names are not established, as they were published without a description.[6]
Related Research Articles
Nepenthes is a genus of carnivorous plants, also known as tropical pitcher plants, or monkey cups, in the monotypic family Nepenthaceae. The genus includes about 170 species, and numerous natural and many cultivated hybrids. They are mostly liana-forming plants of the Old World tropics, ranging from South China, Indonesia, Malaysia, and the Philippines; westward to Madagascar and the Seychelles (one); southward to Australia (four) and New Caledonia (one); and northward to India (one) and Sri Lanka (one). The greatest diversity occurs on Borneo, Sumatra, and the Philippines, with many endemic species. Many are plants of hot, humid, lowland areas, but the majority are tropical montane plants, receiving warm days but cool to cold, humid nights year round. A few are considered tropical alpine, with cool days and nights near freezing. The name "monkey cups" refers to the fact that monkeys were once thought to drink rainwater from the pitchers.
Nepenthes albomarginata is a tropical pitcher plant native to Borneo, Peninsular Malaysia, and Sumatra.
Nepenthes rafflesiana, or Raffles' pitcher-plant, is a species of tropical pitcher plant. It has a very wide distribution covering Borneo, Sumatra, Peninsular Malaysia, and Singapore. Nepenthes rafflesiana is extremely variable, with numerous forms and varieties described. In Borneo alone, there are at least three distinct varieties. The giant form of this species produces enormous pitchers rivalling those of N. rajah in size.
Nepenthes ampullaria is a very distinctive and widespread species of tropical pitcher plant, present in Borneo, the Maluku Islands, New Guinea, Peninsular Malaysia, Singapore, Sumatra, and Thailand.
Nepenthes gracilis, or the slender pitcher-plant, is a common lowland pitcher plant that is widespread in the Sunda region. It has been recorded from Borneo, Cambodia, Peninsular Malaysia, Singapore, Sulawesi, Sumatra, and Thailand. The species has a wide altitudinal distribution of 0 to 1100 m above sea level, although most populations are found below 100 m and plants are rare above 1000 m. Despite being a widespread plant, natural hybrids between N. gracilis and other species are quite rare.
Nepenthes mirabilis, or the common swamp pitcher-plant and tropical pitcher plant, is a carnivorous plant species. By far the most widespread of all Nepenthes, its range covers continental Southeast Asia and all major islands of the Malay Archipelago, stretching from China in the north to Australia in the south. The species exhibits great variability throughout its range. One of the more notable varieties, N. mirabilis var. echinostoma, is endemic to Brunei and Sarawak and possesses an extremely wide peristome.
Nepenthes lowii, or Low's pitcher-plant, is a tropical pitcher plant endemic to Borneo. It is named after Hugh Low, who discovered it on Mount Kinabalu. This species is perhaps the most unusual in the genus, being characterised by its strongly constricted upper pitchers, which bear a greatly reduced peristome and a reflexed lid with numerous bristles on its lower surface.
Nepenthes fusca, or the dusky pitcher-plant, is a tropical pitcher plant endemic to Borneo. It is found throughout a wide altitudinal range and is almost always epiphytic in nature, primarily growing in mossy forest.
Nepenthes × alisaputrana, or the leopard pitcher-plant, is a hybrid of two well-known Nepenthes pitcher plant species: N. burbidgeae and N. rajah. The plant is confined to Mount Kinabalu in Sabah, Borneo.
Dr. Charles M. Clarke is an ecologist and botanist specialising in the carnivorous plant genus Nepenthes, for which he is regarded as a world authority. Clarke has an honours degree in Botany from Monash University in Melbourne, and a Ph.D. in Ecosystem management at the University of New England, in Armidale, New South Wales.
Nepenthes hurrelliana is a tropical pitcher plant endemic to Borneo, where it has been recorded from northern Sarawak, southwestern Sabah, and Brunei. It is of putative hybrid origin; its two original parent species are thought to be N. fusca and N. veitchii. A thick indumentum of rusty-brown hairs covers the entire plant, a characteristic presumably inherited from the latter.
Nepenthes muluensis, or the Mulu pitcher-plant, is a tropical pitcher plant endemic to Borneo. It grows in highland habitats at elevations of 1700 to 2400 m above sea level.
Nepenthes eustachya is a tropical pitcher plant endemic to Sumatra, where it grows from sea level to an elevation of 1600 m. The specific epithet eustachya, formed from the Greek words eu (true) and stachys (spike), refers to the racemose structure of the inflorescence.
Nepenthes inermis is a tropical pitcher plant endemic to the Indonesian island of Sumatra. The specific epithet inermis is Latin for "unarmed" and refers to the upper pitchers of this species, which are unique in that they completely lack a peristome.
Nepenthes × hookeriana, or Hooker's pitcher-plant, is a common natural hybrid involving N. ampullaria and N. rafflesiana. It was originally described as a species.
Colobopsis schmitzi, synonym Camponotus schmitzi, is a species of ant native to Borneo, which is commonly known as the diving ant, swimming ant or pitcher-plant ant, due to their habit of diving into the digestive fluids of their plant host Nepenthes bicalcarata. They are endemic to the island of Borneo.
Nepenthes of Borneo is a monograph by Charles Clarke on the tropical pitcher plants of Borneo. It was first published in 1997 by Natural History Publications (Borneo), and reprinted in 2006. Clarke describes it as "primarily an ecological monograph".
Pitcher-Plants of Borneo is a monograph by Anthea Phillipps and Anthony Lamb on the tropical pitcher plants of Borneo. It was first published in 1996 by Natural History Publications (Borneo), in association with the Royal Botanic Gardens, Kew and the Malaysian Nature Society. An updated and much expanded second edition was published in 2008 as Pitcher Plants of Borneo, with Ch'ien Lee as co-author.
Nepenthes hemsleyana is a tropical pitcher plant endemic to Borneo, where it grows in peat swamp forest and heath forest below 200 m above sea level.
1 2 3 4 5 6 7 8 9 10 11 Bazile, V., J.A. Moran, G. Le Moguédec, D.J. Marshall & L. Gaume 2012. A carnivorous plant fed by its ant symbiont: a unique multi-faceted nutritional mutualism. PLoS ONE7(5): e36179. doi:10.1371/journal.pone.0036179
1 2 Bauer, U., C.J. Clemente, T. Renner & W. Federle 2012. Form follows function: morphological diversification and alternative trapping strategies in carnivorous Nepenthes pitcher plants. Journal of Evolutionary Biology25(1): 90–102. doi:10.1111/j.1420-9101.2011.02406.x
1 2 3 Moran, J.A., B.J. Hawkins, B.E. Gowen & S.L. Robbins 2010. Ion fluxes across the pitcher walls of three Bornean Nepenthes pitcher plant species: flux rates and gland distribution patterns reflect nitrogen sequestration strategies. Journal of Experimental Botany61(5): 1365–1374. doi:10.1093/jxb/erq004
↑ Shelford, R. 1916. A Naturalist in Borneo. T. Fisher Unwin, London.
↑ Cresswell, J.E. 2000. Resource input and the community structure of larval infaunas of an eastern tropical pitcher plant Nepenthes bicalcarata. Ecological Entomology25(3): 362–366. doi:10.1046/j.1365-2311.2000.00267.x
↑ Merbach, M.A., G. Zizka, B. Fiala, U. Maschwitz & W.E. Booth 2000. Carnivory and myrmecophytism—a contradiction? Studies on Nepenthes bicalcarata Hook.f. and its ants. Tagungsband gtö 2000 13. Jahrestagung der Deutschen Gesellschaft für Tropenökologie 1–3. March 2000 in Würzburg Lehrstuhl für Tierökologie und Tropenbiologie Universität Würzburg. p. 106.
↑ Burbidge, F.W. 1880. The Gardens of the Sun: or, a Naturalist's Journal on the Mountains and in the Forests and Swamps of Borneo and the Sulu Archipelago. John Murray, London.
↑ Beccari, O. 1904. Wanderings in the Great Forests of Borneo. Archibald and Constable, London.
↑ Hölldobler, B. & E.O. Wilson 1990. The Ants. Springer-Verlag, Berlin, Germany.
↑ Clarke, C.M. 1992. The ecology of metazoan communities in Nepenthes pitcher plants in Borneo, with special reference to the community of Nepenthes bicalcarata Hook.f. Ph.D. thesis, University of New England, Armidale, New South Wales. 269 pp.
↑ Clarke, C.M. 1998. Initial colonisation and prey capture in Nepenthes bicalcarata (Nepenthaceae) pitchers in Brunei. Sandakania12: 27–36.
↑ Clarke, C.M. 1998. The aquatic arthropod community of the pitcher plant, Nepenthes bicalcarata (Nepenthaceae) in Brunei. Sandakania11: 55–60.
↑ Clarke, C.M. & R.L. Kitching 1993. The metazoan food webs from six Bornean Nepenthes species. Ecological Entomology18(1): 7–16. doi:10.1111/j.1365-2311.1993.tb01074.x
↑ Clarke, C.M. & R.L. Kitching 1995. Swimming ants and pitcher plants: a unique ant-plant interaction from Borneo. Journal of Tropical Ecology11(4): 589–602. doi:10.1017/S0266467400009160
1 2 3 4 5 6 7 Scharmann, M., D.G. Thornham, T.U. Grafe & W. Federle 2013. A novel type of nutritional ant–plant interaction: ant partners of carnivorous pitcher plants prevent nutrient export by dipteran pitcher infauna. PLoS ONE8(5): e63556. doi:10.1371/journal.pone.0063556
↑ Thompson, J.H. 1981. Reverse animal-plant interactions: the evolution of insectivorous and ant-fed plants. Biological Journal of the Linnean Society16: 147–155.
↑ Bohn, H.F., D.G. Thornham & W. Federle 2012. Ants swimming in pitcher plants: kinematics of aquatic and terrestrial locomotion in Camponotus schmitzi. Journal of Comparative Physiology A198(6): 465–476. doi:10.1007/s00359-012-0723-4
1 2 Rembold, K. 2009. Life in Nepenthes Pitchers. In: McPherson, S.R. Pitcher Plants of the Old World. 2 volumes. Redfern Natural History Productions, Poole. pp. 121–141.
↑ Thornham, D.G., J.M. Smith, T.U. Grafe & W. Federle 2012. Setting the trap: cleaning behaviour of Camponotus schmitzi ants increases long-term capture efficiency of their pitcher plant host, Nepenthes bicalcarata. Functional Ecology26(1): 11–19. doi:10.1111/j.1365-2435.2011.01937.x
↑ Clarke, C.M. 1997. The effects of pitcher dimorphism on the metazoan community of the carnivorous pitcher plant Nepenthes bicalcarata Hook.f.. Malayan Nature Journal50: 149–157.
Benz, M.J., E.V. Gorb & S.N. Gorb 2012. Diversity of the slippery zone microstructure in pitchers of nine carnivorous Nepenthes taxa. Arthropod-Plant Interactions6(1): 147–158. doi:10.1007/s11829-011-9171-2
(in French) Blondeau, G. 2001. Nepenthes bicalcarata. In: Les Plantes Carnivores. De Vecchi, Paris. p.70.
Mayer, V.E., M.E. Frederickson, D. McKey & R. Blatrix 2014. Current issues in the evolutionary ecology of ant–plant symbioses. New Phytologist, published online on 21 January 2014. doi:10.1111/nph.12690
Meimberg, H. & G. Heubl 2006. Introduction of a nuclear marker for phylogenetic analysis of Nepenthaceae. Plant Biology8(6): 831–840. doi:10.1055/s-2006-924676
Meimberg, H., S. Thalhammer, A. Brachmann & G. Heubl 2006. Comparative analysis of a translocated copy of the trnK intron in carnivorous family Nepenthaceae. Molecular Phylogenetics and Evolution39(2): 478–490. doi:10.1016/j.ympev.2005.11.023
Osunkoya, O.O., S.D. Daud & F.L. Wimmer 2008. Longevity, lignin content and construction cost of the assimilatory organs of Nepenthes species. Annals of Botany102(5): 845–853. doi:10.1093/aob/mcn162
Takeuchi, Y., M.M. Salcher, M. Ushio, R. Shimizu-Inatsugi, M.J. Kobayashi, B. Diway, C. von Mering, J. Pernthaler & K.K. Shimizu 2011. In situ enzyme activity in the dissolved and particulate fraction of the fluid from four pitcher plant species of the genus Nepenthes. PLoS ONE6(9): e25144. doi:10.1371/journal.pone.0025144
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.






































{kind=link}
{kind=link}
{kind=link}
{kind=link}