Amniotes
Archosauromorphs
Phytosaurs
| Phytosaurs of the Chinle Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | State | Member | Abundance | Notes | Images |
L. crosbiensis [1] |
|     | ||||
S. adamanensis [1] |
| Known from a single skull | Previously referred to Leptosuchus, actually referable to Smilosuchus. [3] | |||
S. gregorii |
|
| Known from a handful of skulls. | A basal leptosuchomorph. | ||
S. lithodendrorum [3] |
|
| Known from only one skull. | Previously considered synonymous with S. adamanensis. | ||
cf. Paleorhinus [4] | Indeterminate [4] |
|
| |||
M. buceros [5] |
| |||||
M. mccauleyi [5] |
| |||||
M. pristinus [5] |
| |||||
| Pravusuchus [3] | P. hortus |
|
| Three skulls. | A leptosuchomorph phytosaur closely related to Mystriosuchini [3] [6] or Nicrosaurus . [7] | |
| Protome [8] | P. batalaria |
|
| Partial skull and jaw. | A phytosaur originally described as a juvenile Leptosuchus , but probably an unrelated non-leptosuchomorph [8] or mystriosuchin. [7] | |
R. gregorii [9] | A large late-surviving mystriosuchin phytosaur closely related to Machaeroprosopus (formerly known as Pseudopalatus ), [10] and often considered a subgenus of that taxon. [2] [6] [7] | |||||
Suchians
Color key
| Notes Uncertain or tentative taxa are in small text; |
| Suchians of the Chinle Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | State | Member | Abundance | Notes | Images |
| Acaenasuchus [11] | A. geoffreyi [11] |
| Known from a handful of isolated osteoderms. [11] | A controversial suchian based primarily on osteoderms. Some paleontologists have considered Acaenasuchus to be juvenile Desmatosuchus haplocerus scutes, while others considered it a separate genus of small aetosaurs. [11] Most recently it has been reconsidered an aetosauriform related to, but not within, Aetosauria. It is also related to Revueltosaurus (also from the Chinle) and the armored reptile Euscolosuchus from Virginia. [12] |         | |
| Adamanasuchus | A. eisenhardtae |
|
| Osteoderms and skull and postcranial fragments. | A basal desmatosuchine aetosaur related to Calyptosuchus and Scutarx . [13] | |
C. wellesi [15] |
|
| Common, although "non-armor material is comparatively rare". [15] | A basal desmatosuchine aetosaur originally considered a species of Stagonolepis . | ||
D. spurensis [17] |
|
| Numerous osteoderms and other skeletal material | A large desmatosuchin aetosaur which was fairly common in the lower part of the Chinle Formation. Prior to 2008, fossils of D. spurensis were listed as the species D. haplocerus, until it was determined that D. haplocerus was a species name originally based on undiagnostic fossils. [13] | ||
D. smalli [18] |
|
| A single lateral osteoderm. | A desmatosuchin aetosaur which is very rare in the Chinle Formation. One of the youngest known occurrences of the genus Desmatosuchus. [13] | ||
E. okeeffeae |
|
| A six-foot-long shuvosaurid from the Coelophysis quarry of Ghost Ranch. | |||
H. agilis [19] |
| An early-diverging crocodylomorph | ||||
| Kryphioparma [20] | K. caerula |
|
| Osteoderms. | Aetosaur described in 2023, probably related to other typothoracines. [20] | |
P. andressorum [21] |
|
| Osteoderms | A significant number of researchers feel that the North American material referred to the German Paratypothorax species P. andressorum by Hunt and Lucas in 1992 is actually distinct. [21] | ||
Indeterminate [22] |
| Osteoderms and minor postcranial fragments. | A paratypothoracin typothoracine aetosaur known from isolated material similar to more complete specimens from Germany. [23] | |||
P. mccreai [24] |
| An early-diverging crocodylomorph | ||||
P. gracilis [25] |
| A Poposaurid pseudosuchian. | ||||
P. kirkpatricki [26] |
| A rauisuchid. | ||||
R. callenderi [28] |
|
| A suchian pseudosuchian. | |||
R. hunti [29] |
|
| ||||
R. chamaensis |
| Osteoderms and possible ankle and vertebrae remains. | A late-surviving paratypothoracin typothoracine aetosaur. Originally referred to Desmatosuchus, but later reinterpreted as a relative of Paratypothorax based on a phylogenetic analysis performed on the remains by Parker in 2003. [30] New genus names were erected independently by Parker (coining the name Heliocanthus) and Lucas et al. (coining the name Rioarribasuchus), prompting an investigation into reports of unethical taxonomic claim-jumping. | |||
Indeterminate [31] |
|
| Jaw fragment. | Identified from a fossil which has been reinterpreted as an indeterminate "rauisuchian". | ||
| Scutarx [13] | S. deltatylus [13] |
|
| Four partial skeletons including skull fragments. | A large basal desmatosuchine aetosaur based on fossils originally referred to "Stagonolepis" (=Calyptosuchus) wellesi, a close relative. Fossils of Scutarx represent the highest stratigraphic occurrence of Stagonolepis-like aetosaurs in Petrified Forest National Park. | |
S. inexpectatus [32] |
| Following a rediagnosis of the genus and species, only shuvosaurid fossils from the Cooper Canyon Formation are reliably identifiable as Shuvosaurus itself. [33] | ||||
| Stenomyti | S. huangae |
|
| Partial skeleton including a well-preserved skull. | A small aetosaurine aetosaur similar to Aetosaurus . | |
T. coccinarum [34] |
| Common in the Owl Rock and Petrified Forest Members. [34] | A large, wide-bodied typothoracine aetosaur abundant throughout the Chinle Formation. Almost all of the known Chinlean Typothorax fossils are isolated whole or fragmentary osteoderms. [34] | |||
| Tecovasuchus [20] | T. chatterjeei |
|
| Osteoderms. | Previously thought to be more widespread in the formation. [20] | |
V. haydeni [35] |
|
| A large rauisuchid similar to Postosuchus . | |||
Ornithodirans
Prosauropod tracks are present in the Redonda, Sloan Canyon, and Sheep Pen Sandstone formations. Possibly the Rock Point Formation as well. Geographically, the tracks are present in New Mexico. [36]
Theropod tracks have been found in Utah and New Mexico recovered from the Redonda, Sloan Canyon, and Sheep Pen Sandstone formations. [36] Indeterminate theropod remains are stratigraphically present in the Petrified Forest, Bluewater Creek, and Rock Point members of New Mexico. [36]
Color key
| Notes Uncertain or tentative taxa are in small text; |
| Ornithodirans of the Chinle Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | State | Member | Abundance | Notes | Images |
C. arizonensis [36] |
|
| A coelophysine coelophysid |  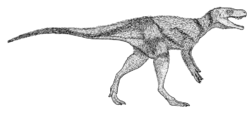      | ||
C. bryansmalli [36] |
|
| A large saurischian alternatively considered a herrerasaurid or a theropod related to Tawa hallae . | |||
C. bauri |
|
| ||||
| C. sp. [38] |
|
| Originally assigned to C. bauri, but likely a different taxon. [38] | |||
C. longicollis [36] |
|
| Now considered a junior synonym of C. bauri. | |||
C. willistoni |
|
| Now considered a junior synonym of C. bauri. | |||
| Dromomeron | D. romeri |
|
| A lagerpetid pterosauromorph found at various Ghost Ranch quarries and elsewhere in the Chinle Formation. | ||
D. chauliodus |
|
| Skull and neck vertebrae fragments | A possible basal theropod with a short skull and enlarged teeth. | ||
E. mcintireae |
|
| Partial mandible, teeth, and wing finger bone | An early-diverging pterosaur | ||
E. baldwini |
|
| A silesaurid dinosauriform closely related to the Polish genus Silesaurus . [40] | |||
K. williamparkeri |
|
| A silesaurid dinosauriform with adaptations for herbivory. | |||
T. hallae |
|
| A probable basal theropod showing a mosaic of features similar to neotheropods and herrerasaurids. | |||
Other archosauromorphs
| Non-crurotarsan Archosauromorphs of the Chinle Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | State | Member | Abundance | Notes | Images |
A. oligos |
| Cervical, dorsal, and caudal vertebrae | A tanystropheid archosauromorph bearing elongated spikes on the vertebral neural spines [42] |       | ||
C. harrisae [43] |
|
| An archosauriform represented only by teeth. [43] It is treated as indeterminate archosauriform remains by Irmis in 2005. [43] It was originally thought to be an ornithischian dinosaur. | |||
| Doswellia [44] | cf. D. kaltenbachi |
|
| Osteoderms, vertebrae, rib and hip fragments. | A doswelliid archosauriform. | |
| Puercosuchus [45] | P. traverorum |
|
| Two monodominant bonebeds. | A carnivorous azendohsaurid archosauromorph. | |
| Rugarhynchos [46] | R. sixmilensis |
|
| Skull and postcranial fragments. | A doswelliid archosauriform. Originally described as a species of Doswellia , [47] but subsequently transferred to a separate genus. | |
| Syntomiprosopus [48] | S. sucherorum |
| Two to four individuals from a single quarry. [48] | A short-faced archosauriform, possibly an unusual early-diverging crocodylomorph. Appears convergent with some Late Cretaceous notosuchians. [48] | ||
| Tanystropheidae [49] | Indeterminate |
|
| Vertebrae | Moderately-sized tanystropheid vertebrae from the Hayden Quarry, likely representing a new taxon closely related to Langobardisaurus and Tanytrachelos . [49] | |
| cf. Tanystropheus [42] | Indeterminate |
|
| Cervical vertebrae | Small tanystropheid neck vertebrae similar to Tanystropheus. [42] | |
|
| A small tanystropheid represented by several hundred fossil specimens. | ||||
T. murryi [51] |
|
| An unknown amniote represented by scattered teeth formerly believed to be from an ornithischian dinosaur. [51] Later discoveries of similar teeth in pseudosuchians meant that these could no longer be regarded as anything more specific than some kind of archosauriform. [51] | |||
T. buettneri [53] |
|
| ||||
T. dornorum [54] |
|
| ||||
T. jacobsi [55] |
| |||||
T. phasmalophos [56] |
|
| ||||
V. campi [57] |
|
| A strange aquatic carnivorous archosauriform, represented by both articulated skeletons and scattered elements like osteoderms and vertebrae. [57] | |||
Other amniotes
| Miscellaneous amniotes of the Chinle Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | State | Member | Abundance | Notes | Images |
A. rectori [59] |
| Known only from a partial skull. [59] | A strange neodiapsid whose bones were heavily ornamented "with subtriangular knobs... running the length of the bones." Even these ornamentations were ornamented "with additional grooves." It is too bizarre to be currently classified as anything more than a probable diapsid. [59] |  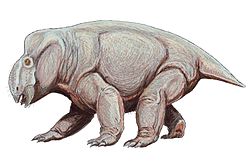 | ||
| Ancistronychus [60] | A. paradoxus |
|
| Manual unguals (hand claws). | A drepanosaur related to Drepanosaurus. | |
| Avicranium [61] | A. renestoi |
|
| Skull and neck vertebrae. | A drepanosaur with a toothless skull and a flexible neck owing to the heterocoelous (saddle-shaped) articular surfaces of the vertebral centrae. The neural spines are anteroposteriorly short and strongly anterodorsally inclined. These features closely resemble those of Drepanosaurus . | |
C. obscurus [62] |
|
| Known only from a jaw fragment and some isolated teeth. [62] | Originally believed to be a fish, Colognathus was a strange amniote with distinctive fluted teeth. [62] | ||
| Dolabrosaurus | D. aquatilis |
|
| Articulated vertebral and limb material. | A drepanosaur. | |
| Drepanosaurus [63] | D. unguicaudatus |
|
| A drepanosaur. | ||
| Fabanychus [64] | F. monos |
|
| A drepanosaur. | ||
K. venetus |
|
| Partial dentaries. | A non-mammalian eucynodont. | ||
K. colberti [66] |
| Known only from two teeth. [66] | A possible cynodont. Although they share some similarities with cynodont teeth, the teeth of Kraterokheirodon are very distinctive and can't be confidently referred to a known amniote group. [66] | |||
| Microzemiotes [67] | M. sonselaensis |
|
| Partial left dentary with three teeth | A diapsid reptile of uncertain affinities. | |
| Palacrodon [68] | P. parkeri [68] |
|
| Jaw fragments. | A diapsid reptile of uncertain phylogenetic placement with unusual broadened teeth. More complete fossils of this genus from the Fremouw Formation of Antarctica suggest that it was an arboreal saurian. [69] | |
P. hesternus [70] |
| Known from several hundred remains, but very rare outside of the highly concentrated Placerias Quarry. [70] | A placeriine stahleckeriid dicynodont. | |||
| Skybalonyx [71] | S. skapter |
|
| Manual unguals (hand claws). | A drepanosaur which likely had a burrowing lifestyle. | |
U. schneideri [73] |
|
| Known in Chinle from only a single tooth. The presence of venom channels is consistent with other known Uatchitodon specimens, although the Chinle specimen's channels are unique in being "completely enclosed under the surface of the crown." [74] | A reptile of unknown affinities, probably a carnivorous archosauromorph with venomous capabilities. | ||
| Whitakersaurus [75] | W. bermani |
|
| A rhynchocephalian found at Ghost Ranch. | ||
| Leptopleuronine procolophonids | Indeterminate |
|
| An incomplete skull (specimen MNA V9953) [76] | ||
|
| Two skulls (specimens MCZ 9312 and 9313) [77] | ||||
Sphenodontia sp. [39] | Indeterminate |
|
| A dentary. | ||
Testudinata sp. [39] | Indeterminate |
|
| Parts of carapaces, cervical osteoderms, a skull roof fragment, left pelvic girdle, a caudal vertebra, a possible squamosal | A terrestrial stem-turtle. | |






