
Eoraptor is a genus of small, lightly built, basal sauropodomorph dinosaur. One of the earliest-known dinosaurs and one of the earliest members of the sauropod family, it lived approximately 231 to 228 million years ago, during the Late Triassic in Western Gondwana, in the region that is now northwestern Argentina. The type and only species, Eoraptor lunensis, was first described in 1993, and is known from an almost complete and well-preserved skeleton and several fragmentary ones. Eoraptor had multiple tooth shapes, which suggests that it was omnivorous. Eoraptor was 1.5 feet (0.46 m) tall and 3 feet (0.91 m) long.

Staurikosaurus is a genus of herrerasaurid dinosaur from the Late Triassic of Brazil, found in the Santa Maria Formation.

Herrerasauridae is a family of carnivorous dinosaurs, possibly basal to either theropods or even all of saurischians, or even their own branching from Dracohors, separate from Dinosauria altogether. They are among the oldest known dinosaurs, first appearing in the fossil record around 233.23 million years ago, before becoming extinct by the end of the Carnian stage. Herrerasaurids were relatively small-sized dinosaurs, normally no more than 4 metres (13 ft) long, although the holotype specimen of "Frenguellisaurus ischigualastensis" is thought to have reached around 6 meters long. The best known representatives of this group are from South America, where they were first discovered in the 1930s in relation to Staurikosaurus and 1960s in relation to Herrerasaurus. A nearly complete skeleton of Herrerasaurus ischigualastensis was discovered in the Ischigualasto Formation in San Juan, Argentina, in 1988. Less complete possible herrerasaurids have been found in North America and Africa, and they may have inhabited other continents as well.

Herrerasaurus is likely a genus of saurischian dinosaur from the Late Triassic period. This genus was one of the earliest dinosaurs from the fossil record. Its name means "Herrera's lizard", after the rancher who discovered the first specimen in 1958 in South America. All known fossils of this carnivore have been discovered in the Ischigualasto Formation of Carnian age in northwestern Argentina. The type species, Herrerasaurus ischigualastensis, was described by Osvaldo Reig in 1963 and is the only species assigned to the genus. Ischisaurus and Frenguellisaurus are synonyms.

Agnosphitys is a genus of dinosauriform that lived during the Late Triassic. It contains only one species, the type species A. cromhallensis. Its remains include an ilium, maxilla, astragalus and humerus, which date variously from the Norian and Rhaetian stages of the Late Triassic, or possibly as late as the Hettangian stage of the Early Jurassic. The fissure fill at Avon, of which Agnosphitys was probably recovered from, was a sinkhole formed by the dissolution of Lower Carboniferous limestones.
Spondylosoma is a genus of avemetatarsalian archosaur belonging to the clade Aphanosauria from the late Ladinian-age Middle Triassic Lower Santa Maria Formation in Paleorrota Geopark, Brazil.
Caseosaurus is a genus of saurischian dinosaur that lived approximately 221.5 to 212 million years ago during the latter part of the Triassic Period in what is now Texas, in North America. It was a small, lightly-built, bipedal, ground-dwelling carnivore, and could grow up to 2 m (6.6 ft) long.
Spinosuchus is an extinct genus of trilophosaurid allokotosaur from the Late Triassic of Texas, southern United States. It has been assigned to a variety of groups over its history, from coelophysid dinosaur to pseudosuchian to uncertain theropod dinosaur and to Proterosuchidae. This uncertainty is not unusual, given that it was only known from a poorly preserved, wall-mounted, partial vertebral column of an animal that lived in a time of diverse, poorly known reptile groups. However, newly collected material and recent phylogenetic studies of early archosauromorphs suggest that it represents an advanced trilophosaurid very closely related to Trilophosaurus.

Poposaurus is an extinct genus of pseudosuchian archosaur from the Late Triassic of the southwestern United States. It belongs to the clade Poposauroidea, an unusual group of Triassic pseudosuchians that includes sail-backed, beaked, and aquatic forms. Fossils have been found in Wyoming, Utah, Arizona, and Texas. Except for the skull, most parts of the skeleton are known. The type species, P. gracilis, was described and named by Maurice Goldsmith Mehl in 1915. A second species, P. langstoni, was originally the type species of the genus Lythrosuchus. Since it was first described, Poposaurus has been variously classified as a dinosaur, a phytosaur, and a "rauisuchian".

Dinosauromorpha is a clade of avemetatarsalians that includes the Dinosauria (dinosaurs) and some of their close relatives. It was originally defined to include dinosauriforms and lagerpetids, with later formulations specifically excluding pterosaurs from the group. Birds are the only dinosauromorphs which survive to the present day.

Saurosuchus is an extinct genus of large loricatan pseudosuchian archosaurs that lived in South America during the Late Triassic period. It was a heavy, ground-dwelling, quadrupedal carnivore, likely being the apex predator in the Ischigualasto Formation.

Dromomeron is a genus of lagerpetid avemetatarsalian which lived around 220 to 211.9 ± 0.7 million years ago. The genus contains species known from Late Triassic-age rocks of the Southwestern United States and northwestern Argentina. It is described as most closely related to the earlier Lagerpeton of Argentina, but was found among remains of true dinosaurs like Chindesaurus, indicating that the first dinosaurs did not immediately replace related groups.

Tawa is a genus of possible basal theropod dinosaurs from the Late Triassic period. The fossil remains of Tawa hallae, the type and only species were found in the Hayden Quarry of Ghost Ranch, New Mexico, US. Its discovery alongside the relatives of Coelophysis and Herrerasaurus supports the hypothesis that the earliest dinosaurs arose in Gondwana during the early Late Triassic period in what is now South America, and radiated from there around the globe. The specific name honours Ruth Hall, founder of the Ghost Ranch Museum of Paleontology.
Lagerpetidae is a family of basal avemetatarsalians. Though traditionally considered the earliest-diverging dinosauromorphs, fossils described in 2020 suggest that lagerpetids may instead be pterosauromorphs. Lagerpetid fossils are known from the Triassic of Argentina, Arizona, Brazil, Madagascar, New Mexico, and Texas. They were typically small, although some lagerpetids, like Dromomeron gigas and a specimen from the Santa Rosa Formation attributed to Dromomeron sp., were able to get quite large. Lagerpetid fossils are rare; the most common finds are bones of the hindlimbs, which possessed a number of unique features.

Eodromaeus is an extinct genus of probable basal theropod dinosaurs from the Late Triassic of Argentina. Like many other of the earliest-known dinosaurs, it hails from the Carnian-age Ischigualasto Formation, within the Ischigualasto-Villa Unión Basin of northwestern Argentina. Upon its discovery, it was argued to be one of the oldest true theropods, supplanting its contemporary Eoraptor, which was reinterpreted as a basal sauropodomorph.
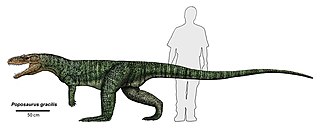
Poposauroidea is a clade of advanced pseudosuchians. It includes poposaurids, shuvosaurids, ctenosauriscids, and other unusual pseudosuchians such as Qianosuchus and Lotosaurus. It excludes most large predatory quadrupedal "rauisuchians" such as rauisuchids and "prestosuchids". Those reptiles are now allied with crocodylomorphs in a clade known as Loricata, which is the sister taxon to the poposauroids in the clade Paracrocodylomorpha. Although it was first formally defined in 2007, the name "Poposauroidea" has been used for many years. The group has been referred to as Poposauridae by some authors, although this name is often used more narrowly to refer to the family that includes Poposaurus and its close relatives.

Daemonosaurus is an extinct genus of possible theropod dinosaur from the Late Triassic of New Mexico. The only known fossil is a skull and neck fragments from deposits of the latest Triassic Chinle Formation at Ghost Ranch. Daemonosaurus was an unusual dinosaur with a short skull and large, fang-like teeth. It lived alongside early neotheropods such as Coelophysis, which would have been among the most common dinosaurs by the end of the Triassic. However, Daemonosaurus retains several plesiomorphic ("primitive") traits of the snout, and it likely lies outside the clade Neotheropoda. It may be considered a late-surviving basal theropod or non-theropod basal saurischian, possibly allied to other early predatory dinosaurs such as herrerasaurids or Tawa.

Buriolestes is a genus of early sauropodomorph dinosaurs from the Late Triassic Santa Maria Formation of the Paraná Basin in southern Brazil. It contains a single species, B. schultzi, named in 2016. The type specimen was found alongside a specimen of the lagerpetid dinosauromorph Ixalerpeton.

Nhandumirim is a genus of basal sauropodomorph dinosaur from the Carnian age of Late Triassic Brazil. It is currently considered a saturnaliid sauropodomorph. The type and only species, Nhandumirim waldsangae, is known from a single immature specimen including vertebrae, a chevron, pelvic material, and a hindlimb found in the Santa Maria Formation in Rio Grande do Sul.

Gnathovorax is a genus of herrerasaurid saurischian dinosaur from the Santa Maria Formation in Rio Grande do Sul, Brazil. The type and only species is Gnathovorax cabreirai, described by Pacheco et al. in 2019.
























