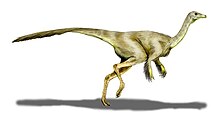
Gallimimus is a genus of theropod dinosaur that lived in what is now Mongolia during the Late Cretaceous period, about seventy million years ago (mya). Several fossils in various stages of growth were discovered by Polish-Mongolian expeditions in the Gobi Desert of Mongolia during the 1960s; a large skeleton discovered in this region was made the holotype specimen of the new genus and species Gallimimus bullatus in 1972. The generic name means "chicken mimic", referring to the similarities between its neck vertebrae and those of the Galliformes. The specific name is derived from bulla, a golden capsule worn by Roman youth, in reference to a bulbous structure at the base of the skull of Gallimimus. At the time it was named, the fossils of Gallimimus represented the most complete and best preserved ornithomimid material yet discovered, and the genus remains one of the best known members of the group.

Oviraptor is a genus of oviraptorid dinosaur that lived in Asia during the Late Cretaceous period. The first remains were collected from the Djadokhta Formation of Mongolia in 1923 during a paleontological expedition led by Roy Chapman Andrews, and in the following year the genus and type species Oviraptor philoceratops were named by Henry Fairfield Osborn. The genus name refers to the initial thought of egg-stealing habits, and the specific name was intended to reinforce this view indicating a preference over ceratopsian eggs. Despite the fact that numerous specimens have been referred to the genus, Oviraptor is only known from a single partial skeleton regarded as the holotype, as well as a nest of about fifteen eggs and several small fragments from a juvenile.

Anserimimus is a genus of ornithomimid theropod dinosaur, from the Late Cretaceous Period of what is now Mongolia. It was a lanky, fast-running animal, possibly an omnivore. From what fossils are known, it probably closely resembled other ornithomimids, except for its more powerful forelimbs.

Ornithomimosauria are theropod dinosaurs which bore a superficial resemblance to the modern-day ostrich. They were fast, omnivorous or herbivorous dinosaurs from the Cretaceous Period of Laurasia, as well as Africa and possibly Australia. The group first appeared in the Early Cretaceous and persisted until the Late Cretaceous. Primitive members of the group include Nqwebasaurus, Pelecanimimus, Shenzhousaurus, Hexing and Deinocheirus, the arms of which reached 2.4 m (8 feet) in length. More advanced species, members of the family Ornithomimidae, include Gallimimus, Struthiomimus, and Ornithomimus. Some paleontologists, like Paul Sereno, consider the enigmatic alvarezsaurids to be close relatives of the ornithomimosaurs and place them together in the superfamily Ornithomimoidea.

Ornithomimus is a genus of ornithomimid theropod dinosaurs from the Campanian and Maastrichtian ages of Late Cretaceous Western North America. Ornithomimus was a swift, bipedal dinosaur which fossil evidence indicates was covered in feathers and equipped with a small toothless beak that may indicate an omnivorous diet. It is usually classified into two species: the type species, Ornithomimus velox, and a referred species, Ornithomimus edmontonicus. O. velox was named in 1890 by Othniel Charles Marsh on the basis of a foot and partial hand from the Denver Formation of Colorado. Another seventeen species have been named since then, though almost all of them have been subsequently assigned to new genera or shown to be not directly related to Ornithomimus velox. The best material of species still considered part of the genus has been found in Alberta, representing the species O. edmontonicus, known from several skeletons from the Horseshoe Canyon Formation. Additional species and specimens from other formations are sometimes classified as Ornithomimus, such as Ornithomimus samueli from the earlier Dinosaur Park Formation.

Dromiceiomimus is a genus of ornithomimid theropod from the Late Cretaceous of Alberta, Canada. The type species, D. brevitertius, is considered a synonym of Ornithomimus edmontonicus by some authors, while others consider it a distinct and valid taxon. It was a small ornithomimid that weighed about 135 kilograms (298 lb).

Deinocheirus is a genus of large ornithomimosaur that lived during the Late Cretaceous around 70 million years ago. In 1965, a pair of large arms, shoulder girdles, and a few other bones of a new dinosaur were first discovered in the Nemegt Formation of Mongolia. In 1970, this specimen became the holotype of the only species within the genus, Deinocheirus mirificus; the genus name is Greek for "horrible hand". No further remains were discovered for almost fifty years, and its nature remained a mystery. Two more complete specimens were described in 2014, which shed light on many aspects of the animal. Parts of these new specimens had been looted from Mongolia some years before, but were repatriated in 2014.

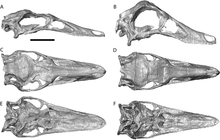
Garudimimus is a genus of ornithomimosaur that lived in Asia during the Late Cretaceous. The genus is known from a single specimen found in 1981 by a Soviet-Mongolian paleontological expedition in the Bayan Shireh Formation and formally described in the same year by Rinchen Barsbold; the only species is Garudimimus brevipes. Several interpretations about the anatomical traits of Garudimimus were made in posterior examinations of the specimen, but most of them were criticized during its comprehensive redescription in 2005. Extensive undescribed ornithomimosaur remains at the type locality of Garudimimus may represent additional specimens of the genus.

Archaeornithomimus is a genus of ornithomimosaurian theropod dinosaur that lived in Asia during the Late Cretaceous period, around 96 million years ago in the Iren Dabasu Formation.
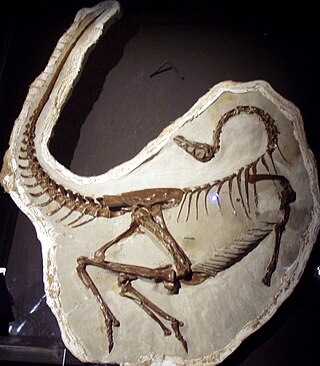
Ornithomimidae is an extinct family of theropod dinosaurs which bore a superficial resemblance to modern ostriches. Ornithomimids were fast, omnivorous or herbivorous dinosaurs known mainly from the Late Cretaceous Period of Laurasia, though they have also been reported from the Lower Cretaceous Wonthaggi Formation of Australia.
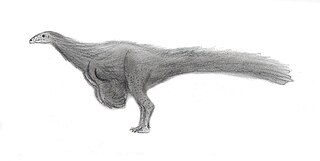
Nqwebasaurus is a basal coelurosaur and is the basal-most member of the coelurosaurian clade Ornithomimosauria from the Early Cretaceous of South Africa. The name Nqwebasaurus is derived from the Xhosa word Nqweba which is the local name for the Kirkwood district, and thwazi is ancient Xhosa for "fast runner". Currently it is the oldest coelurosaur in Africa and shows that basal coelurosaurian dinosaurs inhabited Gondwana 50 million years earlier than previously thought. The type specimen of Nqwebasaurus was discovered by William J. de Klerk who is affiliated with the Albany Museum in Grahamstown. It is the only fossil of its species found to date and was found in the Kirkwood Formation of the Uitenhage Group. Nqwebasaurus has the unofficial nickname "Kirky", due to being found in the Kirkwood.

Kinnareemimus is an extinct genus of ornithomimosaurian theropod dinosaur that was discovered in the Early Cretaceous Sao Khua Formation in what is now Thailand. The genus contains only the type species, K. khonkaenensis. The specific epithet is after Khon Kaen Province, which is in northeastern Thailand, where the remains of the animal were dicovered.

The Nemegt Formation is a geological formation in the Gobi Desert of Mongolia, dating to the Late Cretaceous. The formation consists of river channel sediments and contains fossils of fish, turtles, crocodilians, and a diverse fauna of dinosaurs, including birds.
Deinocheiridae is an extinct family of ornithomimosaurian dinosaurs, living in Asia and the Americas from the Albian until the Maastrichtian. The family was originally named by Halszka Osmólska and Roniewicz in 1970, including only the type genus Deinocheirus. In a 2014 study by Yuong-Nam Lee and colleagues and published in the journal Nature, it was found that Deinocheiridae was a valid family. Lee et al. found that based on a new phylogenetic analysis including the recently discovered complete skeletons of Deinocheirus, the type genus, as well as Garudimimus and Beishanlong, could be placed as a successive group, with Beishanlong as the most primitive and Deinocheirus as most derived. The family Garudimimidae, named in 1981 by Rinchen Barsbold, is now a junior synonym of Deinocheiridae as the latter family includes the type genus of the former. The group existed from 115 to 69 million years ago, with Beishanlong living from 115 to 100 mya, Garudimimus living from 98 to 83 mya, and Deinocheirus living from 71 to 69 mya. Other genera included are Paraxenisaurus, and possibly Harpymimus and Hexing.

Qiupalong is an extinct genus of ornithomimosaurian theropod that was discovered in the Late Cretaceous Qiupa Formation of Henan, China. The genus contains a single species, Q. henanensis, the specific epithet for which was named for the province of Henan. Uniquely, Qiupalong is one of the few Late Cretaceous non-avian dinosaurs known from both Asia and Laramidia. Specimens from Russia and Alberta have been referred to the genus without being assigned to the type species.
"Dryosaurus" grandis is a dubious species of ornithomimosaur dinosaur known from remains found in the Arundel Formation of Maryland.

This timeline of oviraptorosaur research is a chronological listing of events in the history of paleontology focused on the oviraptorosaurs, a group of beaked, bird-like theropod dinosaurs. The early history of oviraptorosaur paleontology is characterized by taxonomic confusion due to the unusual characteristics of these dinosaurs. When initially described in 1924 Oviraptor itself was thought to be a member of the Ornithomimidae, popularly known as the "ostrich" dinosaurs, because both taxa share toothless beaks. Early caenagnathid oviraptorosaur discoveries like Caenagnathus itself were also incorrectly classified at the time, having been misidentified as birds.
This timeline of ornithomimosaur research is a chronological listing of events in the history of paleontology focused on the ornithomimosaurs, a group of bird-like theropods popularly known as the ostrich dinosaurs. Although fragmentary, probable, ornithomimosaur fossils had been described as far back as the 1860s, the first ornithomimosaur to be recognized as belonging to a new family distinct from other theropods was Ornithomimus velox, described by Othniel Charles Marsh in 1890. Thus the ornithomimid ornithomimosaurs were one of the first major Mesozoic theropod groups to be recognized in the fossil record. The description of a second ornithomimosaur genus did not happen until nearly 30 years later, when Henry Fairfield Osborn described Struthiomimus in 1917. Later in the 20th century, significant ornithomimosaur discoveries began occurring in Asia. The first was a bonebed of "Ornithomimus" asiaticus found at Iren Debasu. More Asian discoveries took place even later in the 20th century, including the disembodied arms of Deinocheirus mirificus and the new genus Gallimimus bullatus. The formal naming of the Ornithomimosauria itself was performed by Rinchen Barsbold in 1976.

Rativates is a genus of ornithomimid theropod dinosaur from the Dinosaur Park Formation of Alberta. The type species is Rativates evadens.

Aepyornithomimus is a genus of ornithomimid theropod dinosaur from the Late Cretaceous Djadokhta Formation in Mongolia. It lived in the Campanian, around 75 million years ago, when the area is thought to have been a desert. The type and only species is A. tugrikinensis.