The lateral geniculate nucleus is a relay center in the thalamus for the visual pathway. It receives a major sensory input from the retina. The LGN is the main central connection for the optic nerve to the occipital lobe, particularly the primary visual cortex. In humans, each LGN has six layers of neurons alternating with optic fibers.

The lateral lemniscus is a tract of axons in the brainstem that carries information about sound from the cochlear nucleus to various brainstem nuclei and ultimately the contralateral inferior colliculus of the midbrain. Three distinct, primarily inhibitory, cellular groups are located interspersed within these fibers, and are thus named the nuclei of the lateral lemniscus.
The pretectal area, or pretectum, is a midbrain structure composed of seven nuclei and comprises part of the subcortical visual system. Through reciprocal bilateral projections from the retina, it is involved primarily in mediating behavioral responses to acute changes in ambient light such as the pupillary light reflex, the optokinetic reflex, and temporary changes to the circadian rhythm. In addition to the pretectum's role in the visual system, the anterior pretectal nucleus has been found to mediate somatosensory and nociceptive information.

The reticular formation is a set of interconnected nuclei that are located throughout the brainstem. The reticular formation is not anatomically well defined because it includes neurons located in different parts of the brain. The neurons of the reticular formation make up a complex set of networks in the core of the brainstem that extend from the upper part of the midbrain to the lower part of the medulla oblongata. The reticular formation includes ascending pathways to the cortex in the ascending reticular activating system (ARAS) and descending pathways to the spinal cord via the reticulospinal tracts of the descending reticular formation.

The thalamic reticular nucleus is part of the ventral thalamus that forms a capsule around the thalamus laterally. However, recent evidence from mice and fish question this statement and define it as dorsal thalamic structure. It is separated from the thalamus by the external medullary lamina. Reticular cells are GABAergic, and have discoid dendritic arbors in the plane of the nucleus.

The dentate nucleus is a cluster of neurons, or nerve cells, in the central nervous system that has a dentate – tooth-like or serrated – edge. It is located within the deep white matter of each cerebellar hemisphere, and it is the largest single structure linking the cerebellum to the rest of the brain. It is the largest and most lateral, or farthest from the midline, of the four pairs of deep cerebellar nuclei, the others being the fastigial nucleus and the globose and emboliform nuclei which together are referred to as the interposed nucleus. The dentate nucleus is responsible for the planning, initiation and control of voluntary movements. The dorsal region of the dentate nucleus contains output channels involved in motor function, which is the movement of skeletal muscle, while the ventral region contains output channels involved in nonmotor function, such as conscious thought and visuospatial function.

The tegmentum is a general area within the brainstem. The tegmentum is the ventral part of the midbrain and the tectum is the dorsal part of the midbrain. It is located between the ventricular system and distinctive basal or ventral structures at each level. It forms the floor of the midbrain (mesencephalon) whereas the tectum forms the ceiling. It is a multisynaptic network of neurons that is involved in many subconscious homeostatic and reflexive pathways. It is a motor center that relays inhibitory signals to the thalamus and basal nuclei preventing unwanted body movement. The tegmentum area includes various different structures, such as the "rostral (=frontal/cranial/oral) end of the reticular formation, several nuclei controlling eye movements, the periaqueductal gray matter, the red nucleus, the substantia nigra, and the ventral tegmental area".

The nucleus raphe pallidus receives afferent connections from the periaqueductal gray, the Paraventricular nucleus of hypothalamus, central nucleus of the amygdala, lateral hypothalamic area, and parvocellular reticular nucleus.
Cerebellar peduncles connect the cerebellum to the brain stem. There are six cerebellar peduncles in total, three on each side:
The reticulotegmental nucleus, tegmental pontine reticular nucleus is an area within the floor of the midbrain. This area is known to affect the cerebellum with its axonal projections.
The parvocellular reticular nucleus is part of the brain located dorsolateral to the caudal pontine reticular nucleus.
The caudal pontine reticular nucleus or nucleus reticularis pontis caudalis is composed of gigantocellular neurons.
The gigantocellular nucleus is a subregion of the medullary reticular formation. As the name indicates, is mainly composed of the so-called giant neuronal cells.
The paramedian reticular nucleus sends its connections to the spinal cord in a mostly ipsilateral manner, although there is some decussation.

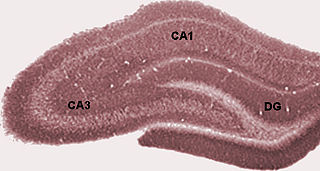
Hippocampus anatomy describes the physical aspects and properties of the hippocampus, a neural structure in the medial temporal lobe of the brain that has a distinctive, curved shape that has been likened to the sea-horse monster of Greek mythology and the ram's horns of Amun in Egyptian mythology. This general layout holds across the full range of mammalian species, from hedgehog to human, although the details vary. For example, in the rat, the two hippocampi look similar to a pair of bananas, joined at the stems. In primate brains, including humans, the portion of the hippocampus near the base of the temporal lobe is much broader than the part at the top. Due to the three-dimensional curvature of this structure, two-dimensional sections such as shown are commonly seen. Neuroimaging pictures can show a number of different shapes, depending on the angle and location of the cut.
The cerebellothalamic tract or the tractus cerebellothalamicus, is part of the superior cerebellar peduncle. It originates in the cerebellar nuclei, crosses completely in the decussation of the superior cerebellar peduncle, bypasses the red nucleus, and terminates in posterior division of ventral lateral nucleus of thalamus. The ventrolateral nucleus has different divisions and distinct connections, mostly with frontal and parietal lobes. The primary motor cortex and premotor cortex get information from the ventrolateral nucleus projections originating in the interposed nucleus and dentate nuclei. Other dentate nucleus projections via thalamic pathway transmit information to prefrontal cortex and posterior parietal cortex. The cerebellum sends thalamocortical projections and in addition may also send connections from the thalamus to association areas serving cognitive and affective functions.
The midbrain reticular formation(MRF) also reticular formation of midbrain, mesencephalic reticular formation, tegmental reticular formation, formatio reticularis (tegmenti) mesencephali) is a structure in the midbrain consisting of the dorsal tegmental nucleus, ventral tegmental nucleus, and cuneiform nucleus. These are also known as the tegmental nuclei.
The rostromedial tegmental nucleus (RMTg), also known as the tail of the ventral tegmental area (tVTA), is a GABAergic nucleus which functions as a "master brake" for the midbrain dopamine system. It is poorly differentiated from the rest of the ventral tegmental area (VTA) and possesses robust functional and structural links to the dopamine pathways. Notably, both acute and chronic exposure to psychostimulants have been shown to induce FosB and ΔFosB expression in the RMTg; no other drug type has been shown to induce these proteins in the RMTg.