
Hepatitis is inflammation of the liver tissue. Some people or animals with hepatitis have no symptoms, whereas others develop yellow discoloration of the skin and whites of the eyes (jaundice), poor appetite, vomiting, tiredness, abdominal pain, and diarrhea. Hepatitis is acute if it resolves within six months, and chronic if it lasts longer than six months. Acute hepatitis can resolve on its own, progress to chronic hepatitis, or (rarely) result in acute liver failure. Chronic hepatitis may progress to scarring of the liver (cirrhosis), liver failure, and liver cancer.

Hepatitis C is an infectious disease caused by the hepatitis C virus (HCV) that primarily affects the liver; it is a type of viral hepatitis. During the initial infection period, people often have mild or no symptoms. Early symptoms can include fever, dark urine, abdominal pain, and yellow tinged skin. The virus persists in the liver, becoming chronic, in about 70% of those initially infected. Early on, chronic infection typically has no symptoms. Over many years however, it often leads to liver disease and occasionally cirrhosis. In some cases, those with cirrhosis will develop serious complications such as liver failure, liver cancer, or dilated blood vessels in the esophagus and stomach.

Hepatocellular carcinoma (HCC) is the most common type of primary liver cancer in adults and is currently the most common cause of death in people with cirrhosis. HCC is the third leading cause of cancer-related deaths worldwide.
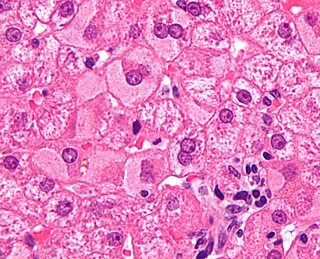
Viral hepatitis is liver inflammation due to a viral infection. It may present in acute form as a recent infection with relatively rapid onset, or in chronic form, typically progressing from a long-lasting asymptomatic condition up to a decompensated hepatic disease and hepatocellular carcinoma (HCC).
The hepatitis C virus (HCV) is a small, enveloped, positive-sense single-stranded RNA virus of the family Flaviviridae. The hepatitis C virus is the cause of hepatitis C and some cancers such as liver cancer and lymphomas in humans.
Human Immunodeficiency Virus (HIV) and Hepatitis C Virus (HCV) co-infection is a multi-faceted, chronic condition that significantly impacts public health. According to the World Health Organization (WHO), 2 to 15% of those infected with HIV are also affected by HCV, increasing their risk of morbidity and mortality due to accelerated liver disease. The burden of co-infection is especially high in certain high-risk groups, such as intravenous drug users and men who have sex with men. These individuals who are HIV-positive are commonly co-infected with HCV due to shared routes of transmission including, but not limited to, exposure to HIV-positive blood, sexual intercourse, and passage of the Hepatitis C virus from mother to infant during childbirth.

C-X-C motif chemokine ligand 10 (CXCL10) also known as Interferon gamma-induced protein 10 (IP-10) or small-inducible cytokine B10 is an 8.7 kDa protein that in humans is encoded by the CXCL10 gene. C-X-C motif chemokine 10 is a small cytokine belonging to the CXC chemokine family.

miR-196 is a non-coding RNA called a microRNA that has been shown to be expressed in humans and mice. miR-196 appears to be a vertebrate specific microRNA and has now been predicted or experimentally confirmed in a wide range of vertebrate species. In many species the miRNA appears to be expressed from intergenic regions in HOX gene clusters. The hairpin precursors are predicted based on base pairing and cross-species conservation—their extents are not known. In this case the mature sequence is excised from the 5' arm of the hairpin.

Liver cancer is cancer that starts in the liver. Liver cancer can be primary or secondary. Liver metastasis is more common than that which starts in the liver. Instances of liver cancer are increasing globally.

Hepatitis B is an infectious disease caused by the Hepatitis B virus (HBV) that affects the liver; it is a type of viral hepatitis. It can cause both acute and chronic infection.

Liver kidney microsomal type 1 antibody (anti-LKM1) is an autoantibody associated with autoimmune hepatitis (AIH). Specifically, its presence in AIH defines type 2 AIH, although it has been proposed that anti-liver cytosol type 1 autoantibody without detectable anti-LKM1 can be seen in type 2 AIH. It is one of the several subtypes of anti–liver-kidney microsome antibodies that are known. The frequent association of anti-LKM-1 antibodies and hepatitis C virus (HCV) infections and the probable existence of an infection-associated autoimmune form of anti-LKM-1-associated hepatitis, requiring a different therapeutic strategy, necessitate the exact determination of anti-LKM-1 specificities.

HBx is a hepatitis B viral protein. It is 154 amino acids long and interferes with transcription, signal transduction, cell cycle progress, protein degradation, apoptosis and chromosomal stability in the host. It forms a heterodimeric complex with its cellular target protein, and this interaction dysregulates centrosome dynamics and mitotic spindle formation. It interacts with DDB1 redirecting the ubiquitin ligase activity of the CUL4-DDB1 E3 complexes, which are intimately involved in the intracellular regulation of DNA replication and repair, transcription and signal transduction.

Hepatitis B virus (HBV) is a partially double-stranded DNA virus, a species of the genus Orthohepadnavirus and a member of the Hepadnaviridae family of viruses. This virus causes the disease hepatitis B.

In molecular biology, mir-221 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.

Interferon lambda 3 encodes the IFNL3 protein. IFNL3 was formerly named IL28B, but the Human Genome Organization Gene Nomenclature Committee renamed this gene in 2013 while assigning a name to the then newly discovered IFNL4 gene. Together with IFNL1 and IFNL2, these genes lie in a cluster on chromosomal region 19q13. IFNL3 shares ~96% amino-acid identity with IFNL2, ~80% identity with IFNL1 and ~30% identity with IFNL4.
In molecular biology mir-198 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.
Infections of the hepatitis C virus (HCV) in children and pregnant women are less understood than those in other adults. Worldwide, the prevalence of HCV infection in pregnant women and children has been estimated to 1-8% and 0.05-5% respectively. The vertical transmission rate has been estimated to be 3-5% and there is a high rate of spontaneous clearance (25-50%) in the children. Higher rates have been reported for both vertical transmission. and prevalence in children (15%).
Christian Bréchot is a French physician and scientist who has been serving as president of the Global Virus Network (GVN) since 2017. He previously served as president of the Institut Pasteur from 2013 until 2017 and as chief executive officer of the French National Institute for Health and Medical Research (INSERM) from 2001 to 2007.
Interferon lambda 4 is one of the most recently discovered human genes and the newest addition to the interferon lambda protein family. This gene encodes the IFNL4 protein, which is involved in immune response to viral infection.

Bulevirtide, sold under the brand name Hepcludex, is an antiviral medication for the treatment of chronic hepatitis D.
