
Prognathodon is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Clidastes. Prognathodon has been recovered from deposits ranging in age from the Campanian to the Maastrichtian in the Middle East, Europe, New Zealand, and North America.

Chimaerasuchus is an extinct genus of Chinese crocodyliform from the Early Cretaceous Wulong Formation. The four teeth in the very tip of its short snout gave it a "bucktoothed" appearance. Due its multicusped teeth and marked heterodonty, it is believed to have been an herbivore. Chimaerasuchus was originally discovered in the 1960s but not identified as a crocodyliform until 1995, instead thought to possibly be a multituberculate mammal. It is highly unusual, as only two other crocodyliforms have displayed any characteristics resembling its adaptations to herbivory.
Platypterygius is a historically paraphyletic genus of platypterygiine ichthyosaur from the Cretaceous period. It was historically used as a wastebasket taxon, and most species within Platypterygius likely are undiagnostic at the genus or species level, or represent distinct genera, even being argued as invalid. While fossils referred to Platypterygius have been found throughout different continents, the holotype specimen was found in Germany.
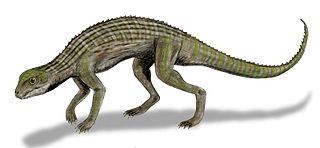
Adamantinasuchus is an extinct genus of notosuchian crocodylomorph from and named after the Late Cretaceous Adamantina Formation of Brazil. It is known from only one fossil, holotype UFRJ-DG 107-R, collected by William Nava. The fossil consists of a partial skull, fragmentary limb bones and a few broken vertebrae, and was found 25 kilometres (16 mi) southwest of the town of Marilia, near a reservoir dam. Adamantinasuchus was approximately 60 centimetres (24 in) long from nose to tail, and would have only weighed a few kilograms.

Plesiotylosaurus, meaning "near Tylosaurus", is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Prognathodon. The genus contains one species, Plesiotylosaurus crassidens, recovered from deposits of Middle Maastrichtian age in the Moreno Formation in California.

Peirosaurus is an extinct genus of peirosaurid crocodylomorph known from the Late Cretaceous period of Minas Gerais, southern Brazil. It contains a single species, Peirosaurus torminni. It is the type genus of the family Peirosauridae.

Xixiasaurus is a genus of troodontid dinosaur that lived during the Late Cretaceous Period in what is now China. The only known specimen was discovered in Xixia County, Henan Province, in central China, and became the holotype of the new genus and species Xixiasaurus henanensis in 2010. The names refer to the areas of discovery, and can be translated as "Henan Xixia lizard". The specimen consists of an almost complete skull, part of the lower jaw, and teeth, as well as a partial right forelimb.

Athabascasaurus is an extinct genus of platypterygiine ophthalmosaurid ichthyosaur known from Alberta, Canada.

Ymeria is an extinct genus of early stem tetrapod from the Devonian of Greenland. Of the two other genera of stem tetrapods from Greenland, Acanthostega and Ichthyostega, Ymeria is most closely related to Ichthyostega, though the single known specimen is smaller, the skull about 10 cm in length. A single interclavicle resembles that of Ichthyostega, an indication Ymeria may have resembled this genus in the post-cranial skeleton.
Eremiasaurus is a genus of mosasaurs, an extinct group of marine reptiles. It lived during the Maastrichtian stage of the Late Cretaceous in what is now North Africa. Only one species is known, E. heterodontus, described in 2012 from two remarkably complete fossil specimens discovered in the Ouled Abdoun Basin, Morocco. This site is known to have delivered a significant number of other related mosasaurs.

Jianchangosaurus is a genus of therizinosaurian dinosaur that lived approximately 126 million years ago during the early part of the Cretaceous Period from the Yixian Formation in what is now China. The type specimen, a juvenile, was discovered in Jianchang County, in the western part of Liaoning Province. It was described in 2013 by a team of palaeontologists from the Henan Museum, who determined that it was among the most basal therizinosaurs.

Pervushovisaurus is a genus of platypterygiine ichthyosaur from the Late Cretaceous (Cenomanian) of the Saratov region in western Russia, the La Penthiève Beds of France and the Cambridge area of the UK.
Muiscasaurus is an extinct genus of ophthalmosaurid ichthyosaur that lived in modern Colombia during the Early Cretaceous. The only known species is the type Muiscasaurus catheti.

Wahasuchus is an extinct genus of engimatic mesoeucrocodylian, likely a neosuchian, of the Middle Campanian age found in the Quseir Formation, Egypt. First described in 2018, Wahasuchus is known mostly from fragmentary remains representing multiple individuals. Given its incomplete nature, it is not entirely clear what its closest relatives are, though features of the skull including its generally flattened morphology akin to that of modern crocodiles suggests it was part of the clade Neosuchia. However it bears no close resemblance to any of the early Cretaceous forms known from northern Africa nor the contemporary taxa of Europe, suggesting that it might have been part of a unique radiation endemic to Africa. The genus currently only contains a single species, Wahasuchus egyptensis.

Arisierpeton is an extinct genus of synapsids from the Early Permian Garber Formation of Richards Spur, Oklahoma. It contains a single species, Arisierpeton simplex.

Thalassodraco is an extinct genus of ophthalmosaurid ichthyosaur from the Late Jurassic (Tithonian) Kimmeridge Clay Formation of England. The type species, T. etchesi, was named in 2020, with the epithet in honour of the discoverer of the holotype, Steve Etches.

Dongnanosuchus is an extinct monotypic genus of orientalosuchin crocodilian known from the middle to late Eocene Youganwo Formation of China. Like other members of Orientalosuchina, Dongnanosuchus was a comparably small-bodied animal with a short, rounded snout that shares characteristics with both early alligatoroids and crocodylids, rendering the precise placement of the clade uncertain. It contains a single species, Dongnanosuchus hsui, which lived during the latter part of the Eocene in what is now the Maoming Basin of China, which at the time featured subtropical forests and a prominent lake that would be the foundation of the local oil shale. This environment was shared by the tomistomine Maomingosuchus and similar cohabitation between Maomingosuchus and orientalosuchins is known from similar sites across East Asia.

Jiangxisuchus is an extinct genus of crocodylian that lived during the Late Cretaceous, likely Maastrichtian, in what is now China. At the time of its description in 2019 it was proposed to be a basal member of Crocodyloidea. However, another concurrent 2019 study recovered Jiangxisuchus instead placed it in the clade Orientalosuchina, which were proposed to be early alligatoroids. The classification of Jiangxisuchus has since then remained in flux. Like other orientalosuchins, Jiangxisuchus was a small to medium-sized animal with a short, blunt snout. The genus is monotypic, containing only the species Jiangxisuchus nankangensis.
Barrosasuchus is a genus of peirosaurid notosuchian from the Santonian of Argentina and part of the extensive peirosaurid record of Late Cretaceous Patagonia. It contains one species, Barrosasuchus neuquenianus. B. neuquenianus is known from an almost complete skull and the majority of the articulated postcranial skeleton, making it the best preserved Patagonian peirosaurid.

Monquirasaurus is an extinct genus of giant short-necked pliosaurs who lived during the Early Cretaceous (Aptian) in what is now Colombia. One species is known, M. boyacensis, described in 2021 from an almost complete fossil skeleton, discovered in 1977 in the town of Villa de Leyva, located in Boyacá. Published descriptions of the holotype specimen estimate that it should reach a total size approaching 9–11 m (30–36 ft) in length and weighing 9–14 metric tons, making Monquirasaurus a large representative of the pliosaurids.










