
Ichthyosauria is an order of large extinct marine reptiles sometimes referred to as "ichthyosaurs", although the term is also used for wider clades in which the order resides.

Temnodontosaurus is an extinct genus of ichthyosaur from the Early Jurassic period. They lived between 200 and 175 million years ago (Hettangian-Toarcian) in what is now Western Europe and possibly other countries including Switzerland and Chile. It lived in the deeper areas of the open ocean. University of Bristol paleontologist Jeremy Martin described the genus Temnodontosaurus as "one of the most ecologically disparate genera of ichthyosaurs," although the number of valid Temnodontosaurus species has varied over the years.

Stenopterygius is an extinct genus of thunnosaur ichthyosaur known from Europe.

Eurhinosaurus is an extinct genus of ichthyosaur from the Early Jurassic (Toarcian), ranging between 183 and 175 million years. Fossils of the aquatic reptile have been found in Western Europe. They used to live in the deep, open sea area. Eurhinosaurus was a large genus of ichthyosaurs. An adult individual could reach up to 7 metres (23 ft) in length.

Aegirosaurus is an extinct genus of platypterygiine ophthalmosaurid ichthyosaurs known from the late Jurassic and early Cretaceous of Europe. It was originally named as a species of Ichthyosaurus.

Chaohusaurus is an extinct genus of basal ichthyosauriform, depending on definition possibly ichthyosaur, from the Early Triassic of Chaohu and Yuanan, China.
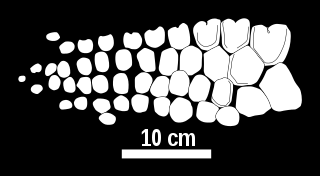
Chacaicosaurus is a genus of neoichthyosaurian ichthyosaur known from the Middle Jurassic of Argentina. The single known specimen of this genus was excavated from the Los Molles Formation in Neuquén Province, and is housed at the Museo Olsacher under the specimen number MOZ 5803. This specimen consists of a skull, forelimb, some vertebrae, and some additional postcranial elements. The genus was named by Marta Fernández in 1994, and contains a single species, Chacaicosaurus cayi, making it the first named distinctive ichthyosaur from the Bajocian stage. It is a medium-sized ichthyosaur with a very long snout, which bears a ridge running along each side. The forelimbs of Chacaicosaurus are small and contain four main digits.

Protoichthyosaurus is a genus of ichthyosaur from the early Jurassic of southern England and possibly Switzerland. Two species are known, P. prostaxalis—the type species, named by Appleby in 1979—and P. applebyi. A third species, P. prosostealis, was named by Appleby, but it was removed from the genus in 2017 due to its similarity to Ichthyosaurus. The genus Protoichthyosaurus was synonymized with Ichthyosaurus by Maisch and Hungerbuhler in 1997, and again by Maisch and Matzke in 2000. However, it was found to be distinct in 2017 by Dean Lomax and colleagues, who separated it from Ichthyosaurus on account of differences in the arrangement and shape of the carpal ossifications, as well as the absence of the fifth digit. The species most likely lived during the Hettangian stage, but may have lived as early as the Rhaetian and as late as the Sinemurian.

Besanosaurus is a extinct genus of Middle Triassic ichthyosaur from Monte San Giorgio of Italy and Switzerland, containing the single species B. leptorhynchus. Besanosaurus was named by Cristiano Dal Sasso and Giovanni Pinna in 1996, based on the nearly complete flattened skeleton BES SC 999, the holotype specimen. This skeleton is preserved across multiple thin rock slabs spanning 3.5 by 4 metres when assembled and took thousands of hours to prepare. Additional specimens from Monte San Giorgio that have previously been considered separate genera, including a partial skull named Mikadocephalus and a well-preserved, largely complete skeleton, have been reinterpreted as additional specimens of Besanosaurus. Putative specimens of Besanosaurus have been discovered in the Norwegian archipelago of Svalbard and Germany, although their attribution to this genus remains disputed.
Callawayia is an extinct genus of ichthyosaur. It contains the species Callawayia neoscapularis.

Guizhouichthyosaurus is an extinct genus of ichthyosaur which is known primarily from the Xiaowa Formation of the lower Carnian stage of the Late Triassic in southwest China. The type species of this genus is Guizhouichthyosaurus tangae, of which multiple skeletons are known. It has been reassigned as a species of the genus Shastasaurus in the past, though it has since been considered distinct. The ichthyosaurs Cymbospondylus asiaticus, named in 2002, and Panjiangsaurus epicharis, named in 2003, are junior synonyms of G. tangae. The genus is also known from the Ladinian-aged Middle Triassic Zhuganpo Formation; additionally, the species "Callawayia" wollongangense may belong to Guizhouichthyosaurus.
Parvinatator, from Latin, “parvus” little and “natator” swimmer, is an extinct genus of small ichthyopterygian marine reptile that lived during the Early to Middle Triassic. Its fossils have been found in British Columbia, Canada.
Suevoleviathan is an extinct genus of primitive ichthyosaur found in the Early Jurassic (Toarcian) of Holzmaden, Germany.

Thaisaurus is an extinct genus of ichthyopterygian marine reptile that lived during the Spathian. Fossils have been found in Thailand.
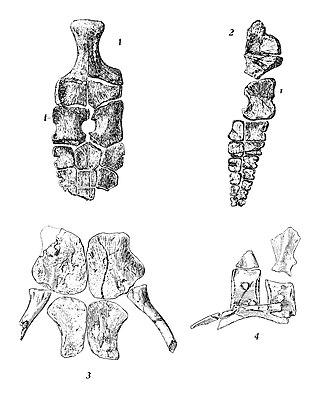
Toretocnemus is an extinct genus of ichthyosaur. Its remains have been found in California, United States, in Triassic layers of the Carnian Hosselkus Limestone.

Wimanius is a genus of ichthyosaur from the Middle Triassic of Switzerland, containing a single species, Wimanius odontopalatus. It was described by Michael Maisch and Andreas Matzke in 1998 based on an incomplete skull from Monte San Giorgio, a mountain on the Swiss-Italian border. Wimanius possesses teeth on its palate, though whether they were located on the palatine or pterygoid is disputed. Other features of Wimanius include a large orbit and jugals with two rami of similar lengths. Different phylogenetic placements of Wimanius have been recovered by different studies, including it being a mixosaurid relative or a merriamosaur, and a monotypic family, Wimaniidae has been named for it. However, its validity has also been questioned, and synonymy with various other genera has been proposed. The only specimen of Wimanius come from the Besano Formation. During the Anisian, this region was a lagoon populated by a wide variety of marine life, including a variety of other ichthyosaurs.

Athabascasaurus is an extinct genus of platypterygiine ophthalmosaurid ichthyosaur known from Alberta, Canada.
Mixosauridae was an early group of ichthyosaurs, living between 247.2 and 235 million years ago, during the Triassic period. Fossils of mixosaurs have been found all over the world: China, Timor, Indonesia, Italy, Germany, Spitsbergen, Switzerland, Svalbard, Canada, Alaska, and Nevada.

This timeline of ichthyosaur research is a chronological listing of events in the history of paleontology focused on the ichthyosauromorphs, a group of secondarily aquatic marine reptiles whose later members superficially resembled dolphins, sharks, or swordfish. Scientists have documented ichthyosaur fossils at least as far back as the late 17th century. At that time, a scholar named Edward Lhuyd published a book on British fossils that misattributed some ichthyosaur vertebrae to actual fishes; their true nature was not recognized until the 19th century. In 1811, a boy named Joseph Anning discovered the first ichthyosaur fossils that would come to be scientifically recognized as such. His sister Mary would later find the rest of its skeleton and would go on to become a respected fossil collector and paleontologist in her own right.

Kyhytysuka is an extinct genus of ophthalmosaurian ichthyosaur from Early Cretaceous Colombia. The animal was previously assigned to the genus Platypterygius, but given its own genus in 2021. Kyhytysuka was a mid-sized ophthalmosaurian with heterodont dentition and several adaptations suggesting that it was a macropredatory vertebrate hunter living in shallow waters. It contains a single species, Kyhytysuka sachicarum.












