
The occipital bone is a cranial dermal bone and the main bone of the occiput. It is trapezoidal in shape and curved on itself like a shallow dish. The occipital bone overlies the occipital lobes of the cerebrum. At the base of the skull in the occipital bone, there is a large oval opening called the foramen magnum, which allows the passage of the spinal cord.

The greater wing of the sphenoid bone, or alisphenoid, is a bony process of the sphenoid bone, positioned in the skull behind each eye. There is one on each side, extending from the side of the body of the sphenoid and curving upward, laterally, and backward.

Macroplata is an extinct genus of Early Jurassic rhomaleosaurid plesiosaur which grew up to 4.65 metres (15.3 ft) in length. Like other plesiosaurs, Macroplata probably lived on a diet of fish, using its sharp needle-like teeth to catch prey. Its shoulder bones were fairly large, indicating a powerful forward stroke for fast swimming. Macroplata also had a relatively long neck, twice the length of the skull, in contrast to pliosaurs.
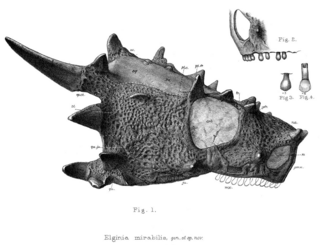
Elginia is an extinct genus of pareiasaurid known from the Late Permian of Scotland and China. It was named for the area around Elgin in Scotland, which has yielded many fossils referred to as the Elgin Reptiles.

Askeptosaurus is an extinct genus of askeptosauroid, a marine reptile from the extinct order Thalattosauria. Askeptosaurus is known from several well-preserved fossils found in Middle Triassic marine strata in what is now Italy and Switzerland.

Shastasaurus is an extinct genus of ichthyosaur from the Late Triassic. Specimens have been found in the United States, Canada, and China.

Anatosuchus is an extinct genus of notosuchian crocodylomorph discovered in Gadoufaoua, Niger, and described by a team of palaeontologists led by the American Paul Sereno in 2003, in the Journal of Vertebrate Paleontology. Its duck-like snout coincidentally makes it resemble a crocoduck, an imagined hybrid animal with the head of a crocodile and the body of a duck.

In human anatomy, the neurocranium, also known as the braincase, brainpan, or brain-pan, is the upper and back part of the skull, which forms a protective case around the brain. In the human skull, the neurocranium includes the calvaria or skullcap. The remainder of the skull is the facial skeleton.

Leptopleuron is an extinct genus of procolophonid that lived in the dry lands during the late Triassic in Elgin of northern Scotland and was the first to be included in the clade of Procolophonidae. First described by English paleontologist and biologist Sir Richard Owen, Leptopleuron is derived from two Greek bases, leptos for "slender" and pleuron for "rib," describing it as having slender ribs. The fossil is also known by a second name, Telerpeton, which is derived from the Greek bases tele for "far off" and herpeton for "reptile." In Scotland, Leptopleuron was found specifically in the Lossiemouth Sandstone Formation. The yellow sandstone it was located in was poorly lithified with wind coming from the southwest. The environment is also described to consist of barchan dunes due to the winds, ranging up to 20 m tall that spread during dry phases into flood plains. Procolophonoids such as Leptopleuron were considered an essential addition to the terrestrial ecosystem during the Triassic.

Chenoprosopus is a genus of extinct cochleosauridae that lived during late Carboniferous and early Permian periods. Two known species of Chenoprosopus are C. milleri and C. lewisi. Chenoprosopus lewisi was described in the basis of a virtually complete skull with maximum skull length of 95 mm. It is significantly smaller than Chenoprosopus milleri and was differentiated from that taxon by Hook (1993) based on sutural patterns of the skull roof. Hook also mentioned the reduced size of the vomerine tusks differentiated C. lewisi from C. milleri, but the different size of these tusks may be different ontogenetic stages of growth. Many of other cochleosaurids from the same time period have an elongated vomer and wide and elongate choana. However, Chenoprosopus is distinguished by its more narrowly pointed snout and separation between the nasal from the maxilla by the broad lacrimal-septomaxilla contact.

Burnetia is an extinct genus of biarmosuchian therapsids in the family Burnetiidae, from the Late Permian of South Africa. Burnetia is known so far from a single holotype skull lacking the lower jaws described by South African paleontologist Robert Broom in 1923. Due to erosion and dorsoventral crushing, features of the skull are hard to interpret. Stutural lines are further distorted by the unusual shape of the skull roof, including many bosses and protuberances.

Plesiopterys is an extinct genus of plesiosaur originating from the Posidonienschiefer of Holzmaden, Germany, and lived during the Early Jurassic period. It is thought to be the sister taxon to all other plesiosauroids including the Plesiosaurus, and is placed outside of the Plesiosauroidea group. Plesiopterys wildi is the one known species within the genus, and is 220 centimeters long, or about 7.2 feet, and its body and skull are both relatively small. It possesses a unique combination of both primitive and derived characters, and is currently displayed at the State Museum of Natural History, Germany.
Callawayia is an extinct genus of ichthyosaur. It contains the species Callawayia neoscapularis.
Wimanius is a genus of ichthyosaur from the Middle Triassic of Switzerland, containing a single species, Wimanius odontopalatus. It was described by Michael Maisch and Andreas Matzke in 1998 based on an incomplete skull from Monte San Giorgio, a mountain on the Swiss-Italian border. Wimanius possesses teeth on its palate, though whether they were located on the palatine or pterygoid is disputed. Other features of Wimanius include a large orbit and jugals with two rami of similar lengths. Different phylogenetic placements of Wimanius have been recovered by different studies, including it being a mixosaurid relative or a merriamosaur, and a monotypic family, Wimaniidae has been named for it. However, its validity has also been questioned, and synonymy with various other genera has been proposed. The only specimen of Wimanius come from the Besano Formation. During the Anisian, this region was a lagoon populated by a wide variety of marine life, including a variety of other ichthyosaurs.

Athabascasaurus is an extinct genus of platypterygiine ophthalmosaurid ichthyosaur known from Alberta, Canada.
Leninia is an extinct genus of basal ophthalmosaurine ichthyosaur known from the late Early Cretaceous of western Russia. Leninia was first named by Valentin Fischer, Maxim S. Arkhangelsky, Gleb N. Uspensky, Ilya M. Stenshin and Pascal Godefroit in 2013 and the type species is Leninia stellans. It was named for Vladimir Lenin, one of the leaders of the Communist Revolution in Russia, but not directlу: the museum where fossils is housed is located within the Lenin Memorial and Lenin school complex in Ulyanovsk; accordingly, the generic name reflects the geohistorical location of the find.
Huskerpeton is an extinct genus of recumbirostran from the Early Permian period. They belong to the order Microsauria, which was established in 1863 by Dawson, and was quickly expanded to include many different small taxa. They lived in what is now Nebraska and Kansas. The holotype of Huskerpeton was uncovered at the Eskridge formation in Nebraska, which is part of how it got its name.

Nebulasaurus is an extinct genus of basal eusauropod dinosaur known from the early Middle Jurassic Zhanghe Formation of Yunnan Province, China. It is known only from the holotype braincase LDRC-v.d.1. A phylogenetic analysis found Nebulasaurus to be a sister taxon to Spinophorosaurus from the Middle Jurassic of Africa. This discovery is significant paleontologically because it represents a clade of basal eusauropods previously unknown from Asia.
This glossary explains technical terms commonly employed in the description of dinosaur body fossils. Besides dinosaur-specific terms, it covers terms with wider usage, when these are of central importance in the study of dinosaurs or when their discussion in the context of dinosaurs is beneficial. The glossary does not cover ichnological and bone histological terms, nor does it cover measurements.

Polymorphodon is an extinct genus of archosauriform reptile from the Middle Triassic of Germany. The only known species is Polymorphodon adorfi, discovered in Lower Keuper deposits at a quarry in Eschenau, Germany. Polymorphodon is notable for its heterodont dentition, with long and conical premaxillary teeth followed by thin maxillary teeth with large serrations. Maxillary teeth near the back of the mouth are short and leaf-shaped, similar to some living and extinct reptiles with a herbivorous or omnivorous diet. This may suggest that Polymorphodon had some reliance on plants in its diet, a rarity among basal archosauriforms, most of which are carnivores.








