
A snake skeleton consists primarily of the skull, vertebrae, and ribs, with only vestigial remnants of the limbs.
Galesaurus is an extinct genus of carnivorous cynodont therapsid that lived between the Induan and the Olenekian stages of the Early Triassic in what is now South Africa. It was incorrectly classified as a dinosaur by Sir Richard Owen in 1859.

Eurhinosaurus is an extinct genus of ichthyosaur from the Early Jurassic (Toarcian), ranging between 183 and 175 million years. Fossils of the aquatic reptile have been found in Western Europe. They used to live in the deep, open sea area. Eurhinosaurus was a large genus of ichthyosaurs. An adult individual could reach up to 7 metres (23 ft) in length.

Askeptosaurus is an extinct genus of askeptosauroid, a marine reptile from the extinct order Thalattosauria. Askeptosaurus is known from several well-preserved fossils found in Middle Triassic marine strata in what is now Italy and Switzerland.

Metoposaurus meaning "front lizard" is an extinct genus of stereospondyl temnospondyl amphibian, known from the Late Triassic of Germany, Italy, Poland, and Portugal. This mostly aquatic animal possessed small, weak limbs, sharp teeth, and a large, flat head. This highly flattened creature mainly fed on fish, which it captured with its wide jaws lined with needle-like teeth. Many Metoposaurus mass graves have been found, probably from creatures that grouped together in drying pools during drought.

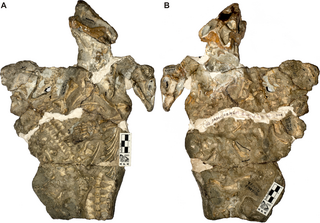
Anatosuchus is an extinct genus of notosuchian crocodylomorph discovered in Gadoufaoua, Niger, and described by a team of palaeontologists led by the American Paul Sereno in 2003, in the Journal of Vertebrate Paleontology. Its duck-like snout coincidentally makes it resemble a crocoduck, an imagined hybrid animal with the head of a crocodile and the body of a duck.

Leptopleuron is an extinct genus of procolophonid that lived in the dry lands during the late Triassic in Elgin of northern Scotland and was the first to be included in the clade of Procolophonidae. First described by English paleontologist and biologist Sir Richard Owen, Leptopleuron is derived from two Greek bases, leptos for "slender" and pleuron for "rib," describing it as having slender ribs. The fossil is also known by a second name, Telerpeton, which is derived from the Greek bases tele for "far off" and herpeton for "reptile." In Scotland, Leptopleuron was found specifically in the Lossiemouth Sandstone Formation. The yellow sandstone it was located in was poorly lithified with wind coming from the southwest. The environment is also described to consist of barchan dunes due to the winds, ranging up to 20 m tall that spread during dry phases into flood plains. Procolophonoids such as Leptopleuron were considered an essential addition to the terrestrial ecosystem during the Triassic.

Mesenosaurus is an extinct genus of amniote. It belongs to the family Varanopidae. This genus includes two species: the type species Mesenosaurus romeri from the middle Permian Mezen River Basin of northern Russia, and Mesenosaurus efremovi from the early Permian (Artinskian) Richards Spur locality. M. romeri’s stratigraphic range is the middle to late Guadalupian while M. efremovi’s stratigraphic range is the Cisuralian.
Parvinatator, from Latin, “parvus” little and “natator” swimmer, is an extinct genus of small ichthyopterygian marine reptile that lived during the Early to Middle Triassic. Its fossils have been found in British Columbia, Canada.

Phantomosaurus is an extinct genus of ichthyosaur that lived during the late Anisian stage of the Middle Triassic. Fossils have been found in southern Germany. It was discovered in 1965 and named in 1997 as a species of Shastasaurus by Sander in the rocks of the Upper Muschelkalk.
Simbirskiasaurus is an extinct genus of ichthyosaur from the Early Cretaceous of Ulyanovsk Oblast, Russia. Its type specimen is YKM 65119, a fragmentary skull and vertebral column.

Polonosuchus is a genus of rauisuchid known from the late Triassic of Poland. It was a huge predator about 5–6 metres in length and, like all rauisuchians, was equipped with a large head of long sharp teeth. The legs were placed almost underneath the body, unlike most reptiles, which would have made it quite fast and a powerful runner. The appearance was very similar to that of the more known Postosuchus, of North America, and shared with the latter the ecological niche of the apex predator.
Australothyris is an extinct genus of basal procolophonomorph parareptile known from the Middle Permian of Tapinocephalus Assemblage Zone, South Africa. The type and only known species is Australothyris smithi. As the most basal member of Procolophonomorpha, Australothyris helped to contextualize the origin of this major parareptile subgroup. It has been used to support the hypotheses that procolophonomorphs originated in Gondwana and ancestrally possess temporal fenestrae, due to its large and fully enclosed temporal fenestra and South African heritage. It also possessed several unique features, including a high tooth number, long postfrontal, small interpterygoid vacuity, and a specialized interaction between the stapes and quadrate.

Athabascasaurus is an extinct genus of platypterygiine ophthalmosaurid ichthyosaur known from Alberta, Canada.

Fodonyx is an extinct genus of rhynchosaur from the middle Triassic epoch of Devon in England. Its fossils were discovered in Otter Sandstone Formation and were first assigned to Rhynchosaurus spenceri. This species was reassigned to its own genus, Fodonyx the holotype of which is EXEMS 60/1985/292, that described by David W. E. Hone and Michael J. Benton in 2008. More recently, one skull was reassigned to the new genus Bentonyx. It is distinguished from other rhynchosaurs by a single autapomorphy, the ventral angling of the paraoccipital processes. In all other rhynchosaurs these processes angle dorsally or are horizontal. It is not known if this conferred any advantage to Fodonyx. Fodonyx was between 40 and 50 cm long.
Dianopachysaurus is an extinct genus of pachypleurosaur known from the lower Middle Triassic of Yunnan Province, southwestern China. It was found in the Middle Triassic Lagerstätte of the Guanling Formation. It was first named by Jun Liu, Olivier Rieppel, Da-Yong Jiang, Jonathan C. Aitchison, Ryosuke Motani, Qi-Yue Zhang, Chang-Yong Zhou and Yuan-Yuan Sun in 2011 and the type species is Dianopachysaurus dingi, thanking a Professor Ding for his help.
Sveltonectes is an extinct genus of platypterygiine ophthalmosaurid ichthyosaurs known from Ul’yanovsk region, western Russia.
Huskerpeton is an extinct genus of recumbirostran from the Early Permian period. They belong to the order Microsauria, which was established in 1863 by Dawson, and was quickly expanded to include many different small taxa. They lived in what is now Nebraska and Kansas. The holotype of Huskerpeton was uncovered at the Eskridge formation in Nebraska, which is part of how it got its name.

Pervushovisaurus is a genus of platypterygiine ichthyosaur from the Late Cretaceous (Cenomanian) of the Saratov region in western Russia, the La Penthiève Beds of France and the Cambridge area of the UK.

Kyhytysuka is an extinct genus of ophthalmosaurian ichthyosaur from Early Cretaceous Colombia. The animal was previously assigned to the genus Platypterygius, but given its own genus in 2021. Kyhytysuka was a mid-sized ophthalmosaurian with heterodont dentition and several adaptations suggesting that it was a macropredatory vertebrate hunter living in shallow waters. It contains a single species, Kyhytysuka sachicarum.







