Fatty acid synthase is a multi-enzyme protein that catalyzes fatty acid synthesis. It is not a single enzyme but a whole enzymatic system composed of two identical 272 kDa multifunctional polypeptides, in which substrates are handed from one functional domain to the next.[1][6][7][8][9]
The fatty acids are synthesized by a series of decarboxylative Claisen condensation reactions from acetyl-CoA and malonyl-CoA. Following each round of elongation the beta keto group is reduced to the fully saturated carbon chain by the sequential action of a ketoreductase (KR), dehydratase (DH), and enoyl reductase (ER). The growing fatty acid chain is carried between these active sites while attached covalently to the phosphopantetheine prosthetic group of an acyl carrier protein (ACP), and is released by the action of a thioesterase (TE) upon reaching a carbon chain length of 16 (palmitic acid).[1]
Classes
There are two principal classes of fatty acid synthases.
Type I systems utilise a single large, multifunctional polypeptide and are common to both animals and fungi (although the structural arrangement of fungal and animal syntheses differ). A Type I fatty acid synthase system is also found in the CMN group of bacteria (corynebacteria, mycobacteria, and nocardia). In these bacteria, the FAS I system produces palmitic acid, and cooperates with the FAS II system to produce a greater diversity of lipid products.[10]
Type II is found in archaea, bacteria and plant plastids, and the mitochondria of animals, including humans, and is characterized by the use of discrete, monofunctional enzymes for fatty acid synthesis. Inhibitors of this pathway (FASII) are being investigated as possible antibiotics.[11]
The mechanism of FAS I and FAS II elongation and reduction is the same, as the domains of the FAS II enzymes are largely homologous to their domain counterparts in FAS I multienzyme polypeptides. However, the differences in the organization of the enzymes - integrated in FAS I, discrete in FAS II - gives rise to many important biochemical differences.[12]
The evolutionary history of fatty acid synthases are very much intertwined with that of polyketide synthases (PKS). Polyketide synthases use a similar mechanism and homologous domains to produce secondary metabolite lipids. Furthermore, polyketide synthases also exhibit a Type I and Type II organization. FAS I in animals is thought to have arisen through modification of PKS I in fungi, whereas FAS I in fungi and the CMN group of bacteria seem to have arisen separately through the fusion of FAS II genes.[10]
Structure
Mammalian FAS consists of a homodimer of two identical protein subunits, in which three catalytic domains in the N-terminal section (-ketoacyl synthase (KS), malonyl/acetyltransferase (MAT), and dehydrase (DH)), are separated by a core region (known as the interdomain) of 600 residues from four C-terminal domains (enoyl reductase (ER), -ketoacyl reductase (KR), acyl carrier protein (ACP) and thioesterase (TE)).[13][14] The interdomain region allows the two monomeric domains to form a dimer.[13]
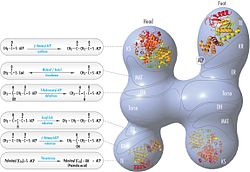
The conventional model for organization of FAS (see the 'head-to-tail' model on the right) is largely based on the observations that the bifunctional reagent 1,3-dibromopropanone (DBP) is able to crosslink the active site cysteine thiol of the KS domain in one FAS monomer with the phosphopantetheine prosthetic group of the ACP domain in the other monomer.[15][16] Complementation analysis of FAS dimers carrying different mutations on each monomer has established that the KS and MAT domains can cooperate with the ACP of either monomer.[17][18] and a reinvestigation of the DBP crosslinking experiments revealed that the KS active site Cys161 thiol could be crosslinked to the ACP 4'-phosphopantetheine thiol of either monomer.[19] In addition, it has been recently reported that a heterodimeric FAS containing only one competent monomer is capable of palmitate synthesis.[20]
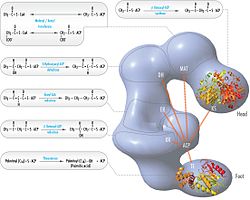
The above observations seemed incompatible with the classical 'head-to-tail' model for FAS organization, and an alternative model has been proposed, predicting that the KS and MAT domains of both monomers lie closer to the center of the FAS dimer, where they can access the ACP of either subunit (see figure on the top right).[21]
A low resolution X-ray crystallography structure of both pig (homodimer)[22] and yeast FAS (heterododecamer)[23] along with a ~6 Å resolution electron cryo-microscopy (cryo-EM) yeast FAS structure [24] have been solved.
Substrate shuttling mechanism
The solved structures of yeast FAS and mammalian FAS show two distinct organizations of highly conserved catalytic domains/enzymes in this multi-enzyme cellular machine. Yeast FAS has a highly efficient rigid barrel-like structure with 6 reaction chambers which synthesize fatty acids independently, while the mammalian FAS has an open flexible structure with only two reaction chambers. However, in both cases the conserved ACP acts as the mobile domain responsible for shuttling the intermediate fatty acid substrates to various catalytic sites. A first direct structural insight into this substrate shuttling mechanism was obtained by cryo-EM analysis, where ACP is observed bound to the various catalytic domains in the barrel-shaped yeast fatty acid synthase.[24] The cryo-EM results suggest that the binding of ACP to various sites is asymmetric and stochastic, as also indicated by computer-simulation studies[25]
FAS revised model with positions of polypeptides, three catalytic domains and their corresponding reactions, visualization by Kosi Gramatikoff. Note that FAS is only active as a homodimer rather than the monomer pictured.
FAS 'head-to-tail' model with positions of polypeptides, three catalytic domains and their corresponding reactions, visualization by Kosi Gramatikoff.
The FASN gene has been investigated as a possible oncogene.[32] FAS is upregulated in breast and gastric cancers, as well as being an indicator of poor prognosis, and so may be worthwhile as a chemotherapeutic target.[33][34][35] FAS inhibitors are therefore an active area of drug discovery research.[36][37][38][39][40]
FAS may also be involved in the production of an endogenous ligand for the nuclear receptor PPARalpha, the target of the fibrate drugs for hyperlipidemia,[41] and is being investigated as a possible drug target for treating the metabolic syndrome.[42]Orlistat which is a gastrointestinal lipase inhibitor also inhibits FAS and has a potential as a medicine for cancer.[43][44]
In some cancer cell lines, this protein has been found to be fused with estrogen receptor alpha (ER-alpha), in which the N-terminus of FAS is fused in-frame with the C-terminus of ER-alpha.[5]
↑ Smith S, Witkowski A, Joshi AK (July 2003). "Structural and functional organization of the animal fatty acid synthase". Progress in Lipid Research. 42 (4): 289–317. doi:10.1016/S0163-7827(02)00067-X. PMID12689621.
↑ Stevens L, Price NC (1999). Fundamentals of enzymology: the cell and molecular biology of catalytic proteins. Oxford [Oxfordshire]: Oxford University Press. ISBN978-0-19-850229-6.
↑ Rangan VS, Joshi AK, Smith S (September 2001). "Mapping the functional topology of the animal fatty acid synthase by mutant complementation in vitro". Biochemistry. 40 (36): 10792–18799. doi:10.1021/bi015535z. PMID11535054.
↑ Anselmi C, Grininger M, Gipson P, Faraldo-Gómez JD (September 2010). "Mechanism of substrate shuttling by the acyl-carrier protein within the fatty acid mega-synthase". Journal of the American Chemical Society. 132 (35): 12357–12364. Bibcode:2010JAChS.13212357A. doi:10.1021/ja103354w. PMID20704262.
↑ Na M, Jang J, Min BS, Lee SJ, Lee MS, Kim BY, Oh WK, Ahn JS (September 2006). "Fatty acid synthase inhibitory activity of acylphloroglucinols isolated from Dryopteris crassirhizoma". Bioorganic & Medicinal Chemistry Letters. 16 (18): 4738–4742. doi:10.1016/j.bmcl.2006.07.018. PMID16870425.
↑ Baron A, Migita T, Tang D, Loda M (January 2004). "Fatty acid synthase: a metabolic oncogene in prostate cancer?". Journal of Cellular Biochemistry. 91 (1): 47–53. doi:10.1002/jcb.10708. PMID14689581. S2CID26175683.
↑ Hunt DA, Lane HM, Zygmont ME, Dervan PA, Hennigar RA (2007). "MRNA stability and overexpression of fatty acid synthase in human breast cancer cell lines". Anticancer Research. 27 (1A): 27–34. PMID17352212.
↑ Gansler TS, Hardman W, Hunt DA, Schaffel S, Hennigar RA (June 1997). "Increased expression of fatty acid synthase (OA-519) in ovarian neoplasms predicts shorter survival". Human Pathology. 28 (6): 686–692. doi:10.1016/S0046-8177(97)90177-5. PMID9191002.
↑ Lu T, Schubert C, Cummings MD, Bignan G, Connolly PJ, Smans K, Ludovici D, Parker MH, Meyer C, Rocaboy C, Alexander R, Grasberger B, De Breucker S, Esser N, Fraiponts E, Gilissen R, Janssens B, Peeters D, Van Nuffel L, Vermeulen P, Bischoff J, Meerpoel L (May 2018). "Design and synthesis of a series of bioavailable fatty acid synthase (FASN) KR domain inhibitors for cancer therapy". Bioorganic & Medicinal Chemistry Letters. 28 (12): 2159–2164. doi:10.1016/j.bmcl.2018.05.014. PMID29779975. S2CID29159508.
↑ Hardwicke MA, Rendina AR, Williams SP, Moore ML, Wang L, Krueger JA, Plant RN, Totoritis RD, Zhang G, Briand J, Burkhart WA, Brown KK, Parrish CA (September 2014). "A human fatty acid synthase inhibitor binds β-ketoacyl reductase in the keto-substrate site". Nature Chemical Biology. 10 (9): 774–779. doi:10.1038/nchembio.1603. PMID25086508.
Ye Q, Chung LW, Li S, Zhau HE (2000). "Identification of a novel FAS/ER-alpha fusion transcript expressed in human cancer cells". Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression. 1493 (3): 373–377. doi:10.1016/s0167-4781(00)00202-5. PMID11018265.
1xkt: Human fatty acid synthase: Structure and substrate selectivity of the thioesterase domain
2cg5: STRUCTURE OF AMINOADIPATE-SEMIALDEHYDE DEHYDROGENASE-PHOSPHOPANTETHEINYL TRANSFERASE IN COMPLEX WITH CYTOSOLIC ACYL CARRIER PROTEIN AND COENZYME A
2jfd: STRUCTURE OF THE MAT DOMAIN OF HUMAN FAS
2jfk: STRUCTURE OF THE MAT DOMAIN OF HUMAN FAS WITH MALONYL-COA
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.