Haplogroup C is believed to have arisen in East Asia[4] some 24,000 years before present. It is a descendant of the haplogroup M. Haplogroup C shares six mutations downstream of the MRCA of haplogroup M with haplogroup Z and five mutations downstream of the MRCA of haplogroup M with other members of haplogroup M8. This macro-haplogroup is known as haplogroup M8'CZ or simply as haplogroup M8.
Distribution
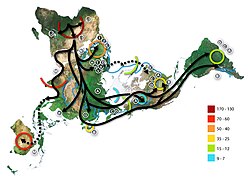
Frequency distribution of mtDNA haplogroup C in EurasiaApproximate geographical distribution of the C1 sub-clades.
In 2010, Icelandic researchers discovered C1e lineage in their home country, estimating an introduction date of year 1700 AD or earlier, indicating a possible introduction during the Viking expeditions to the Americas. A Native American origin for this C1e lineage is likely, but the researchers note that a European or Asian one cannot be ruled out.[10][11][12]
In 2014, a study discovered a new mtDNA subclade C1f from the remains of 3 people found in north-western Russia and dated to 7,500 years ago. It has not been detected in modern populations. The study proposed the hypothesis that the sister C1e and C1f subclades had split early from the most recent common ancestor of the C1 clade and had evolved independently. Subclade C1e had a northern European origin. Iceland was settled by the Vikings 1,130 years ago and they had raided heavily into western Russia, where the sister subclade C1f is now known to have resided. They proposed that both subclades were brought to Iceland through the Vikings, however C1e went extinct on mainland northern Europe due to population turnover and its small representation, and subclade C1f went extinct completely.[13]
In 2015, a study conducted in the Aconcagua mummy identified its mtDNA lineage belongs to the subclade C1bi, which contains 10 distinct mutations from C1b.[14]
This phylogenetic tree of haplogroup C subclades is based on the paper by Mannis van Oven and Manfred Kayser Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation[5] and subsequent published research.
C-T195C! – Ireland, Scotland, England, USA, Hungary (Szeged region[32]), Poland, Belarus,[citation needed] Russia (Russian,[2] Buryat), Turkey, Pakistan (Hazara), India (Jammu and Kashmir), China (Bargut and Mongol in Inner Mongolia, etc.), Korea
C4a1a2 – China
C4a1a2a – China (Han from Ili, Han from Henan, etc.)
C4a1a2b
C4a1a2b1 - China
C4a1a2b2 - Uyghur
C4a1a3 – Bronze Age Irkutsk Oblast (Ust'-Belaya, Khaptsagai, Silinskij, Chastaja Padi), Russian (Kemerovo Oblast[3]), Koryak,[29] Yukaghir,[7][8] Yakut,[8] Evenk (Nyukzha,[8]Chumikan,[29] Nelkan/Dzhigda[29]), Even (Sakkyryyr,[8] Sebjan,[8]Tompo,[8] Markovo,[29] Kamchatka[8]), Udinsk Buryat (Kushun[29]), Todzhin (Toora-Hem,[29] Adir-Kezhig[29]), Altai Kizhi,[2] Iran (Qashqai[33]), Sweden
C4c1b – Chippewa (Trempealeau in Wisconsin),[citation needed] Ottawa or Chippewa (Sault Saint Marie, Chippewa County, Michigan),[citation needed] Canada
C7 – South Korea,[2] China,[45] Taiwan (Hakka), Thailand (Khon Mueang in Chiang Rai Province, Chiang Mai Province, and Lamphun Province[38]), Vietnam (Kinh, Tay, Jarai)
C7a – Han (Beijing, Yunnan, Denver, etc.),[46] Uyghur, Taiwan (Paiwan, Minnan), Lahu, Thailand (incl. Urak Lawoi, Lao Isan in Chaiyaphum Province, Khon Mueang in Lamphun Province, Khon Mueang in Lampang Province, Kaleun in Nakhon Phanom Province, Black Tai in Loei Province, Phuan in Suphan Buri Province[38]), Vietnam (Hani, Yao, Gelao)
C7a1 – China, Mongol (Alxa[16]), Taiwan (Makatao), USA (Han Chinese in Denver), Korea
C7a2 – China,[49]Dai, Laos (Lao in Luang Prabang[38]), Thailand (Khon Mueang in Chiang Mai Province, Khon Mueang in Chiang Rai Province, Lao Isan in Roi Et Province, Phutai in Sakon Nakhon Province, Mon in Nakhon Ratchasima Province[38]), Myanmar (Yangon)
C7a2a – China (Shantou, etc.),[50] Taiwan (Hakka, Makatao, etc.)
12Derenko, Miroslava; Malyarchuk, Boris; Grzybowski, Tomasz; Denisova, Galina; Rogalla, Urszula; Perkova, Maria; Dambueva, Irina; Zakharov, Ilia (2010). "Origin and Post-Glacial Dispersal of Mitochondrial DNA Haplogroups C and D in Northern Asia". PLOS ONE. 5 (12) e15214. Bibcode:2010PLoSO...515214D. doi:10.1371/journal.pone.0015214. ISSN1932-6203. PMC3006427. PMID21203537. More than a half of the northern Asian pool of mtDNA is fragmented into a number of subclades of haplogroups C and D, two of the most frequent haplogroups throughout northern, eastern, central Asia and America. Previous studies have proposed that haplogroups C and D originated around 30–50 kya in eastern Asia, from where they subsequently expanded northwards to southern Siberia, and further deep into northern Asia and the Americas, and westwards along the Steppe Belt extending from Manchuria to Europe [14], [15]. It has been also shown that haplogroups C and D were strongly involved in the late-glacial expansions from southern China to northeastern India [16]. In addition, because of their high frequency and wide distribution, haplogroups C and D most likely participated in all subsequent episodes of putative gene flow in northern Eurasia. These include (i) the Paleolithic colonization of Siberia that is associated with the development of macroblade industries (40–30 kya), (ii) further recolonization and possible replacement of early Siberians by microblade-making human populations from the Lake Baikal, Yenisei River, and Lena River basin regions (20 kya), (iii) appearance of pottery-making Neolithic tradition in the forest-steppe belt of northern Eurasia starting at about 14.5 kya and its expanding into the East European Plane (7 kya), (iv) the Neolithic dispersal of agriculture in eastern Asia, (v) the expansion of the Afanasievo and Andronovo cultures (5–3 kya), and (vi) more recent events of gene flow to eastern and central Europe."
↑Sunna Ebenesersdóttir, Sigríður (2010). "A new subclade of mtDNA haplogroup C1 found in icelanders: Evidence of pre-columbian contact?". American Journal of Physical Anthropology. 144 (1): 92–99. doi:10.1002/ajpa.21419. PMID21069749.
123456789101112Guang‐Lin He, Meng‐Ge Wang, Xing Zou, Hui‐Yuan Yeh, Chang‐Hui Liu, Chao Liu, Gang Chen, and Chuan‐Chao Wang. Extensive ethnolinguistic diversity at the crossroads of North China and South Siberia reflects multiple sources of genetic diversity[J]. J Syst Evol, 2023, 61(1): 230-250.
1234567Starikovskaya, E.B., Sukernik, R.I., Derbeneva, O.A., Volodko, N.V., Ruiz-Pesini, E., Torroni, A., Brown, M.D., Lott, M.T., Hosseini, S.H., Huoponen, K. and Wallace, D.C., "Mitochondrial DNA diversity in indigenous populations of the southern extent of Siberia, and the origins of Native American haplogroups." Annals of Human Genetics 69 (PT 1), 67-89 (2005).
123Marchi, Nina; Hegay, Tatyana; Mennecier, Philippe; Georges, Myriam; Laurent, Romain; Whitten, Mark; Endicott, Philipp; Aldashev, Almaz; Dorzhu, Choduraa; Nasyrova, Firuza; Chichlo, Boris; Ségurel, Laure; Heyer, Evelyne (2017). "Sex-specific genetic diversity is shaped by cultural factors in Inner Asian human populations". American Journal of Physical Anthropology. 162 (4): 627–640. Bibcode:2017AJPA..162..627M. doi:10.1002/ajpa.23151. PMID28158897.
1234567U.A. Perego, N. Angerhofer, M. Pala, et al., "The initial peopling of the Americas: a growing number of founding mitochondrial genomes from Beringia." Genome Res. 20 (9), 1174-1179 (2010).
↑S. Kumar, C. Bellis, M. Zlojutro, et al., "Large scale mitochondrial sequencing in Mexican Americans suggests a reappraisal of Native American origins." BMC Evol. Biol. 11 (1), 293 (2011).
12R.S. Malhi, J.S. Cybulski, R.Y. Tito, et al., "Brief communication: Mitochondrial haplotype C4c confirmed as a founding genome in the Americas." Am. J. Phys. Anthropol. 141 (3), 494-497 (2010).
12345Peng, Min-Sheng; Xu, Weifang; Song, Jiao-Jiao; Chen, Xing; Sulaiman, Xierzhatijiang; Cai, Liuhong; Liu, He-Qun; Wu, Shi-Fang; Gao, Yun; Abdulloevich, Najmudinov Tojiddin; Afanasevna, Manilova Elena; Ibrohimovich, Khudoidodov Behruz; Chen, Xi; Yang, Wei-Kang; Wu, Miao; Li, Gui-Mei; Yang, Xing-Yan; Rakha, Allah; Yao, Yong-Gang; Upur, Halmurat; Zhang, Ya-Ping (2018). "Mitochondrial genomes uncover the maternal history of the Pamir populations". European Journal of Human Genetics. 26 (1): 124–136. doi:10.1038/s41431-017-0028-8. PMC5839027. PMID29187735.
123Kilinc, G.M., Kashuba, N., Yaka, R., Sumer, A.P., Yuncu, E., Shergin, D., Ivanov, G.L., Kichigin, D., Pestereva, K., Volkov, D., Mandryka, P., Kharinskii, A., Tishkin, A., Ineshin, E., Kovychev, E., Stepanov, A., Alekseev, A., Fedoseeva, S.A., Somel, M., Jakobsson, M., Krzewinska, M., Stora, J., and Gotherstrom, A., "Investigating Holocene human population history in North Asia using ancient mitogenomes." Sci Rep 8 (1), 8969 (2018)
123456Derenko, M. V.; Grzybowski, T.; Malyarchuk, B. A.; Dambueva, I. K.; Denisova, G. A.; Czarny, J.; Dorzhu, C. M.; Kakpakov, V. T.; Miscicka-Sliwka, D.; Wozniak, M.; Zakharov, I. A. (September 2003). "Diversity of Mitochondrial DNA Lineages in South Siberia". Annals of Human Genetics. 67 (5): 391–411. doi:10.1046/j.1469-1809.2003.00035.x. PMID12940914. S2CID28678003.
↑Klunk, J., Duggan, A.T., Redfern, R., et al., "Genetic resiliency and the Black Death: No apparent loss of mitogenomic diversity due to the Black Death in medieval London and Denmark." Am. J. Phys. Anthropol. (2019).
↑O. A. Derbeneva, E. B. Starikovskaya, N. V. Volodko, D. C. Wallace, and R. I. Sukernik, "Mitochondrial DNA Variation in the Kets and Nganasans and Its Implications for the Initial Peopling of Northern Eurasia." Russian Journal of Genetics, Vol. 38, No. 11, 2002, pp. 1316–1321. Translated from Genetika, Vol. 38, No. 11, 2002, pp. 1554–1560.
↑Malyarchuk,B., Litvinov,A., Derenko,M., Skonieczna,K., Grzybowski,T., Grosheva,A., Shneider,Y., Rychkov,S. and Zhukova,O., "Mitogenomic diversity in Russians and Poles." Forensic Sci Int Genet 30, 51-56 (2017).
↑Jiang,C., Cui,J., Liu,F., Gao,L., Luo,Y., Li,P., Guan,L. and Gao,Y., "Mitochondrial DNA 10609T Promotes Hypoxia-Induced Increase of Intracellular ROS and Is a Risk Factor of High Altitude Polycythemia." PLoS ONE 9 (1), E87775 (2014).
Bermisheva, M. A.; Kutuev, I. A.; Spitsyn, V. A.; Villems, R.; Batyrova, A. Z.; Korshunova, T. Yu.; Khusnutdinova, E. K. (January 2005). "Analysis of Mitochondrial DNA variation in the population of oroks". Russian Journal of Genetics. 41 (1): 66–71. doi:10.1007/PL00022112. S2CID264200417.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.