
The Cochin forest cane turtle, also known as Kavalai forest turtle, forest cane turtle or simply cane turtle, is a rare turtle from the Western Ghats of India. Described in 1912, its type locality is given as "Near Kavalai in the Cochin State Forests, inhabiting dense forest, at an elevation of about 1500 feet above sea level". Only two specimens were found at that time, and no scientist saw this turtle for the next 70 years. It was rediscovered in 1982, and since then a number of specimens have been found and some studies have been conducted about its phylogeny and ecology.
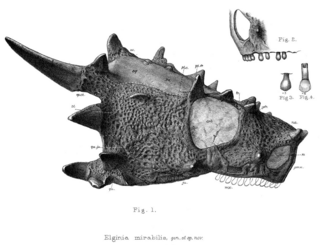
Elginia is an extinct genus of pareiasaurid known from the Late Permian of Scotland and China. It was named for the area around Elgin in Scotland, which has yielded many fossils referred to as the Elgin Reptiles.

Stupendemys is an extinct genus of freshwater side-necked turtle, belonging to the family Podocnemididae. It is the largest freshwater turtle known to have existed, with a carapace over 2 meters long. Its fossils have been found in northern South America, in rocks dating from the Middle Miocene to the very start of the Pliocene, about 13 to 5 million years ago. Male specimens are known to have possessed bony horns growing from the front edges of the shell and the discovery of the fossil of a young adult shows that the carapace of these turtles flattens with age. A fossil skull described in 2021 indicates that Stupendemys was a generalist feeder.

Meiolania is an extinct genus of meiolaniid stem-turtle native to Australasia throughout much of the Cenozoic. Meiolania was a large turtle, with the shell alone ranging from 0.7–2 m in length. Four species are currently recognized, although the validity of two of them is disputed. Meiolania was first described as a species of lizard related to Megalania by Richard Owen towards the end of the 19th century, before the continued discovery of additional fossils solidified its placement as a kind of turtle.

Meiolaniidae is an extinct family of large, probably herbivorous stem-group turtles with heavily armored heads and clubbed tails known from South America and Australasia. Though once believed to be cryptodires, they are not closely related to any living species of turtle, and lie outside crown group Testudines, having diverged from them around the Middle Jurassic. They are best known from the last surviving genus, Meiolania, which lived in Australia from the Miocene until the Pleistocene, and insular species that lived on Lord Howe Island and New Caledonia during the Pleistocene and possibly the Holocene for the latter. Meiolaniids are part of the broader grouping of Meiolaniformes, which contains more primitive turtles species lacking the distinctive morphology of meiolaniids, known from the Early Cretaceous-Paleocene of South America and Australia.

Ninjemys oweni is an extinct large meiolaniid stem-turtle from Pleistocene Queensland and possibly New South Wales (Australia). It overall resembled its relative, Meiolania, save that the largest pair of horns on its head stuck out to the sides, rather than point backwards, the larger scales at the back of its skull and the tail club which is made up of only two tail rings rather than four. With a shell length of approximately 1 m it is a large turtle and among the largest meiolaniids. Ninjemys is primarily known from a well preserved skull and associated tail armor, which were initially thought to have belonged to the giant monitor lizard Megalania.

Niolamia is an extinct genus of South American meiolaniid turtle with a long and complex history. Like its relatives, Niolamia was a robust and heavily armored terrestrial turtle with large, horn like scales covering its head and a tail encased by rings of bone. This heavily armored build may have served the animal during intraspecific combat during courtship, though such encounters likely did not involve the horns and frill, which are thought to serve more of a display function. Scans of the skull reveal that Niolamia likely had a great sense of smell but only low frequency hearing, indicating that these animals communicated more through chemical signals and smells than through sound.

The turtle shell is a shield for the ventral and dorsal parts of turtles, completely enclosing all the vital organs of the turtle and in some cases even the head. It is constructed of modified bony elements such as the ribs, parts of the pelvis and other bones found in most reptiles. The bone of the shell consists of both skeletal and dermal bone, showing that the complete enclosure of the shell likely evolved by including dermal armor into the rib cage.

Acherontemys is an extinct genus of turtle from Eocene sediments in northwestern North America and comprising a single species Acherontemys heckmani. Acherontemys has been placed within the pond turtle superfamily Testudinoidea as part of the clade Pan-Emydidae.
Condorchelys was a genus of stem turtle from Early Jurassic Cañadon Asfalto Formation of Argentina. Condorchelys represents the oldest Jurassic-aged Turtle from South America, with only one species described, Condorchelys antiqua.

Warkalania is an extinct genus of Australian meiolaniid turtle from the Oligocene or early Miocene of Riversleigh, Queensland. While other meiolaniids are known for their elaborate headcrests or long horns, Warkalania only possesses very short horns that form a somewhat continuous ridge across the back of the head. The only known species of this genus, Warkalania carinaminor, is the oldest named meiolaniid turtle of Australia.

Cyamodontoidea is an extinct superfamily of placodont marine reptiles from the Triassic period. It is one of the two main groups of placodonts, the other being Placodontoidea. Cyamodontoids are distinguished from placodontoids by their large shells, formed from fused bony plates called osteoderms and superficially resembling the shells of turtles. Cyamodontoids also have distinctive skulls with narrow, often toothless jaws and wide, flaring temporal regions behind the eyes. Two large temporal openings are positioned at the top of the back of the skull, an arrangement that is known as the euryapsid condition and seen throughout Sauropterygia, the marine reptile group to which placodonts belong. Cyamodontoids are also distinguished by their large crushing teeth, which grow from the palatine bones on the roof of the mouth.
Psephosauriscus is an extinct genus of placodont reptile from the Middle Triassic of Israel and Egypt. It is known from bony armor plates that have been found from Makhtesh Ramon in Israel's Negev desert and Araif en Naqua on Egypt's Sinai Peninsula. The genus was erected in 2002 as a replacement name for several species of the genus Psephosaurus, which was named in 1957. It includes the species P. mosis, P. ramonensis, P. sinaiticus, and a possible fourth species, P. rhombifer. All species, with the exception of P. ramonensis, were once assigned to the genus Psephosaurus. Remains of P. mosis and P. ramonensis were found in Makhtesh Ramon, while P. sinaiticus and P. rhombifer were found in Araif en Naqua.
Chubutemys was an extinct genus of meiolaniform turtle. It lived during the Early Cretaceous of Argentina, around the Albian-Aptian border, within the Puesto La Paloma Member of the Cerro Barcino Formation. It is known from most of the skeleton and carapace, and part of the skull.

Kunbarrasaurus is an extinct genus of small ankylosaurian dinosaur from the Cretaceous of Australia. The genus conatins a single species, K. ieversi.

Basilemys is a large, terrestrial nanhsiungchelyid turtle from the Upper Cretaceous of North and Central America. Basilemys has been found in rocks dating to the Campanian and Maastrichtian subdivisions of the Late Cretaceous and is considered to be the largest terrestrial turtle of its time. In an analysis made by Sukhanov et al. on a nansiunghelyid turtle from the Upper Cretaceous of Mongolia, it was demonstrated that Asian nanhsiungchelyids gave rise to the North American nanhsiungchelyids.
Osteoderms are dermal bone structures that support the upper layer of skin and serve as protection against the elements in a large variety of extinct and extant organisms, especially reptiles. This structure is commonly called "dermal armor" and serves to protect the organism, while also helping with temperature regulation. Osteoderms represent hard tissue components of the integument, making them easy to identify in fossil examination. This dermal armor is found prominently in many lizards. Some early amphibians have this armor, but it is lost in modern species with the exception a ventral plate, called the gastralia.

Meiolaniformes is an extinct clade of stem-group turtles, defined as all taxa more closely related to Meiolania than to Cryptodira and Pleurodira. It is known from the Early Cretaceous to the Holocene of Australia, Oceania and South America. Some Eurasian taxa have been suggested to be part of the group, but this is disputed.
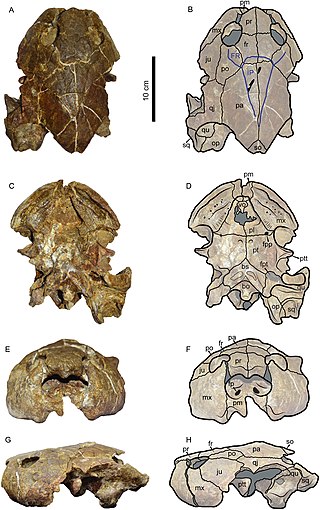
Caninemys is an extinct genus of large freshwater side-necked turtle, belonging to the family Podocnemididae. Its fossils have been found in Brazil and Colombia, in rocks dating back from the middle to late Miocene.

Stegouros is a genus of ankylosaurian dinosaur from the Late Cretaceous Dorotea Formation of southern Chile. The genus contains a single species, Stegouros elengassen, known from a semi-articulated, near-complete skeleton.

