
Megatherium is an extinct genus of ground sloths endemic to South America that lived from the Early Pliocene through the end of the Pleistocene. It is best known for the elephant-sized type species Megatherium americanum, native to the Pampas through southern Bolivia during the Pleistocene. Various other species belonging to the subgenus Pseudomegatherium ranging in size comparable to considerably smaller than M. americanum are known from the Andean region.
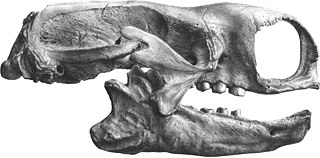
Mylodon is a genus of extinct ground sloth belonging to the family Mylodontidae, known from southern South America. With a total length of 3 to 4 m and a body mass of 1-2 tonnes, it is one of the largest mylodontids.

Ground sloths are a diverse group of extinct sloths in the mammalian superorder Xenarthra. Ground sloths varied widely in size, with the largest, belonging to genera Eremotherium and Megatherium, being around the size of elephants. Ground sloths are a paraphyletic group, as living tree sloths are thought to have evolved from ground sloth ancestors.

Glossotherium is an extinct genus of large mylodontid ground sloths of the subfamily Mylodontinae. It represents one of the best-known members of the family, along with Mylodon and Paramylodon. Reconstructed animals were between 3 and 4 metres long and possibly weighed up to 1,002.6–1,500 kg. The majority of finds of Glossotherium date from the Middle and Upper Pleistocene, around 300,000 to 10,000 years ago, with a few dating older, as far back Pliocene, about 3.3-3 million years ago. The range included large parts of South America, east of the Andes roughly from latitude 20 to 40 degrees south, leaving out the Amazon Basin in the north. In western South America, finds are also documented north of the equator. The animals largely inhabited the open landscapes of the Pampas and northern savanna regions.

Eremotherium is an extinct genus of giant ground sloth in the family Megatheriidae. Eremotherium lived in southern North America, Central America, and northern South America from the Pliocene, around 5.3 million years ago, to the end of the Late Pleistocene, around 10,000 years ago. Eremotherium was one of the largest ground sloths, with a body size comparable to elephants, weighing around 4–6.5 tonnes and measuring about 6 metres (20 ft) long, slightly larger than its close relative Megatherium.

Paramylodon is an extinct genus of ground sloth of the family Mylodontidae endemic to North America during the Pliocene through Pleistocene epochs, living from around ~4.9 Mya–12,000 years ago.

Lestodon is an extinct genus of giant ground sloth native to South America during the Pleistocene epoch. Its fossil remains have been primarily been found in the Pampas and adjacent regions. The largest member of the family Mylodontidae, It is estimated to have weighed 4,100 kilograms. It was a herbivore and primarily fed on the grasses and low-growing plants.
Mylodontinae is an extinct subfamily of ground sloths that lived from the Early Miocene to the Early Holocene epochs.
Lestobradys is an extinct genus of ground sloth, which existed in Uruguay during the Late Miocene period; Huayquerian in the South American land mammal age (SALMA). The type species is L. sprechmanni, found in the Camacho Formation of Uruguay.

Oreomylodon is an extinct genus of ground sloth in the family Mylodontidae, endemic to Ecuador during the Pleistocene. The only species, O. wegneri, was long considered to be either a species or subgenus of Glossotherium or a junior synonym of Glossotherium robustum, but studies of its cranial anatomy published in 2019 have supported Oreomylodon as a valid genus, and suggested it is more closely related to Paramylodon. However, a subsequent analysis published in 2020 again sunk Oreomylodon wegneri into Glossotherium, as a distinct species. It shows adaptations to living in a high-altitude habitat, and its fossils have frequently been unearthed in the Interandean Valles of Ecuador, at elevations of between 2,450 and 3,100 meters.
Megatheriinae is a subfamily of the Megatheriidae, an extinct family of ground sloths that lived from the Middle Miocene to the Early Holocene.

Pseudoprepotherium is an extinct genus of sloths of the family Mylodontidae. It was widespread across northern South America during the Early to Late Miocene epoch around 21 to 5.3 million years ago. Fossils of the animal have been found in Brazil, Venezuela, and Peru. Pseudoprepotherium lived in a tropical climate with a water-rich environment. Their known remains are limited to limb bones, except for a few skulls and teeth. Based on these remains, they were most likely medium to large-sized mylodontid. The genus was described in 1961 and currently contains three species, which were originally assigned to the genus Prepotherium.
Megathericulus is an extinct genus of ground sloths in the Megatheriidae family. It lived during the Middle Miocene, 11-16 Ma in what is now South America. Fossils have been found principally in Argentina, Bolivia, and Peru. It is a smaller representative of the megatheres. Despite being one of the earliest-known members of the family, its dentition structure is associated with homodont teeth belonging to the more modern line of evolution. The genus was scientifically named in 1904. Only one species is currently recognized, Megathericulus patagonicus.

Proeremotherium is an extinct genus of megatheriine ground sloths in the family Megatheriidae. It lived during the Late Miocene and Early Pliocene of what is now Venezuela. So far, two largely complete skulls have been recovered in the Falcón Basin in Venezuela. The finds identify the animals as medium-sized representatives of the Megatheriidae. In the cranial anatomy, Proeremotherium resembles the later and giant Eremotherium. It is therefore assumed that the two ground sloths are directly related to each other.
Archaeomylodon is an extinct genus of mylodontine ground sloths that lived during the Middle Pleistocene of what is now Argentina. It is known so far only from a single skull, which in its dimensions corresponds to those of the giant Lestodon. However, the skull differs from this one by its narrower and higher snout. In addition, the anterior canine teeth, which are usually large in many mylodonts, are greatly reduced. The find comes from the Pampa region of South America and was deposited in about 700,000 years old sediments.
Urumacotherium is an extinct genus of ground sloths of the family Mylodontidae. It lived from the Middle Miocene to the Early Pliocene of what is now Brazil, Peru and Venezuela.
Baraguatherium is an extinct genus of ground sloths of the family Mylodontidae that lived during the Early Miocene of what is now Venezuela. It dates to the Early Miocene, around 20.44 to 15.97 million years ago and represents the oldest representative of its family in the northern part of South America to date. The structure of the teeth suggests that the genus represents a rather basal form within the Mylodontidae. Unlike other mylodonts, which tended to prefer open grasslands, Baraguatherium lived in a riverine, coastal tropical rainforest.
Lakukullus is an extinct genus of nothrotheriid ground sloths that lived during the Middle Miocene around 13.8 to 11.8 million years ago of what is now Bolivia.
Aymaratherium is an extinct genus of nothrotheriid ground sloths that lived during the Late Miocene and Early Pliocene of Bolivia. Fossils of Aymaratherium have been found in the Pomata Ayte locality of the Umala Formation.
Bolivartherium is an extinct genus of mylodontine mylodontid sloth that lived during the Late Miocene and Late Pliocene in what is now Venezuela. Fossils have been found in the Codore and Urumaco Formations of Venezuela.

