This enzyme belongs to the family of transferases, specifically those transferring phosphorus-containing groups (phosphotransferases) with a carboxy group as acceptor. The systematic name of this enzyme class is ATP:butanoate 1-phosphotransferase. This enzyme participates in butyrate metabolism.
The reaction above is a nucleophilic substitution reaction. An electron pair from an oxygen on ADP attacks the phosphorus on butyryl-phosphate, breaking the bond between phosphorus and oxygen to create ATP and butyrate. The arrow-pushing mechanism is shown above.
The reaction can also occur in the reverse direction, as shown below, under certain fermentation conditions.[4][5]
ATP + butyrate ADP + butyryl-phosphate
Structure
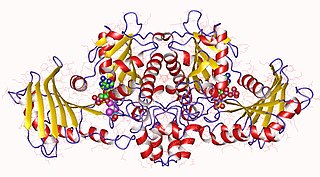
As of 2015, two structures have been solved for this class of enzymes, with PDB accession codes 1SAZ and 1X9J. The study conducted to solve 1SAZ was retracted in 2012 due to fact that the data was used without the permission of the sole custodian.[6]
The investigators of the study that produced the crystallization of 1X9J hypothesized that the enzyme was an octomer formed from dimers.[3] The crystallized form has a radius of 7.5nm which corresponded to a molecular weight of 380kDa. Because a monomer of buk2 is about 43kDa, it was believed that the enzyme itself was either an octomer or a nonamer. Investigators hypothesized that the enzyme was an octomer since most of the proteins within the ASHKA super family form dimers.
Function
Butyrate kinase is active within the human colon.[1] To form butyrate, two molecules of acetyl-CoA are combined and reduced to produce butyryl-CoA. Butyryl CoA is then converted into butyrate through two reactions. The first reaction converts butyryl-CoA to butyryl-phosphate by using the phosphotransbutyrylase enzyme.[2] Butyryl-phosphate is then converted into butyrate by using butyrate kinase and in the process, releases ATP.[7][8]
Because of the significant roles that butyrate plays within cells, it is essential that butyrate kinase is functioning correctly, which can be done through regulation of the enzyme. One study has previously found that butyrate kinase is not regulated by its end-products or other acids such as acetic acid,[11] but more studies need to be conducted to further elucidate the regulation of butyrate kinase.
Disease relevance
As stated in the previous section, butyrate is involved with multiple cellular functions. Because of its involvement with these functions, it is hypothesized that butyrate can act as a protective agent against colon cancer and various inflammatory bowel diseases.[7][9][10][12] Butyrate plays a key role in colon cancer by switching its role concerning cellular proliferation and apoptosis depending on the state and conditions of the cell.[9][10] Butyrate also possesses anti-inflammatory effects to decrease colonic inflammation such as ulcerative colitis.[9] One study specifically identified the transcription factorNF-kB as a target of butyrate to decrease the number of pro-inflammatory cytokines.[7]
Related Research Articles
In enzymology, a 1-phosphatidylinositol 4-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a 5-methyldeoxycytidine-5'-phosphate kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a dephospho-CoA kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a formate kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a glutamate 5-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a glycerate kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a hamamelose kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a N-acetylgalactosamine kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a polynucleotide 5'-hydroxyl-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a protein-histidine pros-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a protein-histidine tele-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a pseudouridine kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a rhamnulokinase is an enzyme that catalyzes the chemical reaction
In enzymology, a scyllo-inosamine 4-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a streptomycin 6-kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a tagatose kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a thiamine-diphosphate kinase is an enzyme involved in thiamine metabolism. It catalyzes the chemical reaction
In enzymology, a thiamine kinase is an enzyme that catalyzes the chemical reaction
In enzymology, a thiamine-phosphate kinase is an enzyme that catalyzes the chemical reaction
In molecular biology, acetate kinase (EC 2.7.2.1), which is predominantly found in micro-organisms, facilitates the production of acetyl-CoA by phosphorylating acetate in the presence of ATP and a divalent cation. Short-chain fatty acids (SCFAs) play a major role in carbon cycle and can be utilized as a source of carbon and energy by bacteria. Salmonella typhimurium propionate kinase (StTdcD) catalyzes reversible transfer of the γ-phosphate of ATP to propionate during l-threonine degradation to propionate. Kinetic analysis revealed that StTdcD possesses broad ligand specificity and could be activated by various SCFAs (propionate>acetate≈butyrate), nucleotides (ATP≈GTP>CTP≈TTP; dATP>dGTP>dCTP) and metal ions (Mg2+≈Mn2+>Co2+). Inhibition of StTdcD by tricarboxylic acid (TCA) cycle intermediates such as citrate, succinate, α-ketoglutarate and malate suggests that the enzyme could be under plausible feedback regulation. Crystal structures of StTdcD bound to PO4 (phosphate), AMP, ATP, Ap4 (adenosine tetraphosphate), GMP, GDP, GTP, CMP and CTP revealed that binding of nucleotide mainly involves hydrophobic interactions with the base moiety and could account for the broad biochemical specificity observed between the enzyme and nucleotides. Modelling and site-directed mutagenesis studies suggest Ala88 to be an important residue involved in determining the rate of catalysis with SCFA substrates. Molecular dynamics simulations on monomeric and dimeric forms of StTdcD revealed plausible open and closed states, and also suggested role for dimerization in stabilizing segment 235-290 involved in interfacial interactions and ligand binding. Observation of an ethylene glycol molecule bound sufficiently close to the γ-phosphate in StTdcD complexes with triphosphate nucleotides supports direct in-line phosphoryl transfer. The enzyme is important in the process of glycolysis, enzyme levels being increased in the presence of excess glucose. The growth of a bacterial mutant lacking acetate kinase has been shown to be inhibited by glucose, suggesting that the enzyme is involved in excretion of excess carbohydrate. A related enzyme, butyrate kinase, facilitates the formation of butyryl-CoA by phosphorylating butyrate in the presence of ATP to form butyryl phosphate.
1 2 Walter KA, Nair RV, Cary JW, Bennett GN, Papoutsakis ET (November 1993). "Sequence and arrangement of two genes of the butyrate-synthesis pathway of Clostridium acetobutylicum ATCC 824". Gene. 134 (1): 107–11. doi:10.1016/0378-1119(93)90182-3. PMID8244020.
1 2 Diao J, Cooper DR, Sanders DA, Hasson MS (June 2003). "Crystallization of butyrate kinase 2 from Thermotoga maritima mediated by vapor diffusion of acetic acid". Acta Crystallographica D. 59 (Pt 6): 1100–2. doi:10.1107/s0907444903007832. PMID12777787.
↑ Rogers, P (1986). "Genetics and biochemistry of Clostridium relevant to development of fermentation processes". Advances in Applied Microbiology Volume 31. pp.1–60. doi:10.1016/s0065-2164(08)70438-6. ISBN9780120026319.{{cite book}}: |journal= ignored (help)
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.