Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of Ascomycota are asexual and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

Polypores are a group of fungi that form large fruiting bodies with pores or tubes on the underside. They are a morphological group of basidiomycetes-like gilled mushrooms and hydnoid fungi, and not all polypores are closely related to each other. Polypores are also called bracket fungi or shelf fungi, and they characteristically produce woody, shelf- or bracket-shaped or occasionally circular fruiting bodies that are called conks. Over one thousand polypore species have been described to science, but a large part of the diversity is still unknown even in relatively well-studied temperate areas.

Gleba is the fleshy spore-bearing inner mass of certain fungi such as the puffball or stinkhorn.

Scleroderma is a genus of fungi, commonly known as earth balls, now known to belong to the Boletales order, in suborder Sclerodermatineae. The best known species are S. citrinum and S. verrucosum. They are found worldwide. Various members of this genus are used as inoculation symbionts to colonize and promote the growth of tree seedlings in nurseries. They are not edible.

Cyathus striatus, commonly known as the fluted bird's nest, is a common saprobic bird's nest fungus with a widespread distribution throughout temperate regions of the world. This fungus resembles a miniature bird's nest with numerous tiny "eggs"; the eggs, or peridioles, are actually lens-shaped bodies that contain spores. C. striatus can be distinguished from most other bird's nest fungi by its hairy exterior and grooved inner walls. Although most frequently found growing on dead wood in open forests, it also grows on wood chip mulch in urban areas. The fruiting bodies are encountered from summer until early winter. The color and size of this species can vary somewhat, but they are typically less than a centimeter wide and tall, and grey or brown in color. Another common name given to C. striatus, splash cups, alludes to the method of spore dispersal: the sides of the cup are angled such that falling drops of water can dislodge the peridioles and eject them from the cup. The specific epithet is derived from the Latin stria, meaning "with fine ridges or grooves".

Crucibulum is a genus in the Nidulariaceae, a family of fungi whose fruiting bodies resemble tiny egg-filled bird's nests. Often called "splash cups", the fruiting bodies are adapted for spore dispersal by using the kinetic energy of falling drops of rain. The "eggs" inside the bird's nests are hard waxy shells containing spores, and tend to stick to whatever nearby herbage they land on, thus increasing the odds of being consumed and dispersed by herbivorous animals. Members of this genus are saprobic, obtaining nutrients from dead organic matter, and are typically found growing on decayed wood and wood debris. The three known Crucibulum species are distinguished from other genera of the Nidulariaceae by their relatively simple funiculus – a cord of hyphae that connects the peridiole to the exterior of the bird's nest.

Nidularia is a genus of nine species of fungi in the family Agaricaceae. Their fruit bodies resemble tiny egg-filled bird nests. The name comes from the Latin nidus meaning nest. The related genus Mycocalia was segregated from Nidularia in 1961 based on differences in the microscopic structure of the peridium.

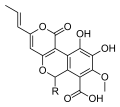
Nidula is a genus of fungi in the family Agaricaceae. Their fruit bodies resemble tiny egg-filled birds' nests, from which they derive their common name "bird's nest fungi". Originally described in 1902, the genus differs from the related genera Cyathus and Crucibulum by the absence of a cord that attaches the eggs to the inside of the fruit body. The life cycle of this genus allows it to reproduce both sexually, with meiosis, and asexually via spores. Species in this genus produce a number of bioactive compounds, including 4-(p-hydroxyphenyl)-2-butanone, a major component of raspberry flavor and insect attractor used in pesticides.
Mycocalia is a genus of fungi in the family Nidulariaceae. Basidiocarps are minute and irregularly spherical. Each produces one or more peridioles which contain the spores and are released from the disintegrating fruit bodies at maturity. Species are usually found growing on herbaceous stems and other plant debris. The genus was originally described in 1961 by British mycologist J.T. Palmer and has a north temperate distribution.

The peridium is the protective layer that encloses a mass of spores in fungi. This outer covering is a distinctive feature of gasteroid fungi.

Cyathus olla also known as the field bird's nest is a species of saprobic fungus in the genus Cyathus of the family Nidulariaceae. The fruit bodies resemble tiny bird's nests filled with "eggs" – spore-containing structures called peridioles. Like other bird's nest fungi, C. olla relies on the force of falling water to dislodge peridioles from fruiting bodies to eject and disperse their spores. The life cycle of this fungus allows it to reproduce both sexually, with meiosis, and asexually via spores. C. olla is a relatively common fungus, with a worldwide distribution. It is the subject of agricultural research to determine its potential as a means to accelerate the breakdown of crop residue, and reduce the population of plant pathogens. The specific epithet is derived from the Latin word olla, meaning "pot".

Cyathus stercoreus, commonly known as the dung-loving bird's nest or the dung bird's nest, is a species of fungus in the genus Cyathus, family Nidulariaceae. Like other species in the Nidulariaceae, the fruiting bodies of C. stercoreus resemble tiny bird's nests filled with eggs. The fruiting bodies are referred to as splash cups, because they are developed to use the force of falling drops of water to dislodge and disperse their spores. The species has a worldwide distribution, and prefers growing on dung, or soil containing dung; the specific epithet is derived from the Latin word stercorarius, meaning "of dung".

Cyathus helenae or Helena's bird's nest is a species of fungus in the genus Cyathus, family Nidulariaceae. Like other members of the Nidulariaceae, C. helenae resembles a tiny bird's nest filled with 'eggs'—spore-containing structures known as peridioles. It was initially described by mycologist Harold Brodie in 1966, who found it growing on mountain scree in Alberta, Canada. C. helenae's life cycle allows it to reproduce both sexually and asexually. One of the smaller species of Cyathus, C. helenae produces a number of chemically unique diterpenoid molecules known as cyathins. The specific epithet of this species was given by Brodie in tribute to his late wife Helen.

A coprophilous fungus is a type of saprobic fungus that grows on animal dung. The hardy spores of coprophilous species are unwittingly consumed by herbivores from vegetation, and are excreted along with the plant matter. The fungi then flourish in the faeces, before releasing their spores to the surrounding area.

Clavaria is a genus of fungi in the family Clavariaceae. Species of Clavaria produce basidiocarps that are either cylindrical to club-shaped or branched and coral-like. They are often grouped with similar-looking species from other genera, when they are collectively known as the clavarioid fungi. All Clavaria species are terrestrial and most are believed to be saprotrophic. In Europe, they are typical of old, mossy, unimproved grassland. In North America and elsewhere, they are more commonly found in woodlands.

The gasteroid fungi are a group of fungi in the Basidiomycota. Species were formerly placed in the obsolete class Gasteromycetes Fr., or the equally obsolete order Gasteromycetales Rea, because they produce spores inside their basidiocarps rather than on an outer surface. However, the class is polyphyletic, as such species—which include puffballs, earthballs, earthstars, stinkhorns, bird's nest fungi, and false truffles—are not closely related to each other. Because they are often studied as a group, it has been convenient to retain the informal (non-taxonomic) name of "gasteroid fungi".
The Nidulariaceae are a family of fungi in the order Agaricales. Commonly known as the bird's nest fungi, their fruiting bodies resemble tiny egg-filled birds' nests. As they are saprobic, feeding on decomposing organic matter, they are often seen growing on decaying wood and in soils enriched with wood chips or bark mulch; they have a widespread distribution in most ecological regions. The five genera within the family, namely, Crucibulum, Cyathus, Mycocalia, Nidula, and Nidularia, are distinguished from each other by differences in morphology and peridiole structure; more recently, phylogenetic analysis and comparison of DNA sequences is guiding new decisions in the taxonomic organization of this family.
Retiperidiolia is a genus of fungi in the family Nidulariaceae. Basidiocarps are typically under 10 mm in diameter and irregularly spherical. Each produces a number of peridioles which contain the spores and are released from the disintegrating fruit bodies at maturity. Species are usually found growing on herbaceous stems and other plant debris. The genus has a tropical distribution. Species were previously referred to Mycocalia, but molecular research, based on cladistic analysis of DNA sequences, found that they were not closely related.

Hexagonia vesparia, sometimes known as the wasp nest polypore, is a bracket fungus in the Polyporaceae family occurring mainly in tropical and coastal regions in Australia, but it has also been recorded in semi arid regions of Australia. The genus name came from the Latin word hexagonus meaning with six angles.




![A peridiole and attached funiculus in cross section
.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key>ol{margin-left:1.3em;margin-top:0}.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key>ul{margin-top:0}.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key li{page-break-inside:avoid;break-inside:avoid-column}@media(min-width:300px){.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key,.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key-wide{column-count:2}.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key-narrow{column-count:1}}@media(min-width:450px){.mw-parser-output figure[typeof="mw:File/Thumb"] .image-key-wide{column-count:3}}
.mw-parser-output .plainlist ol,.mw-parser-output .plainlist ul{line-height:inherit;list-style:none;margin:0;padding:0}.mw-parser-output .plainlist ol li,.mw-parser-output .plainlist ul li{margin-bottom:0}
a: tunica
b: spores
c: basidia
d: purse
e: funicular cord
f: hapteron
g: middle piece Peridiole cross section.svg](http://upload.wikimedia.org/wikipedia/commons/thumb/1/13/Peridiole_cross_section.svg/220px-Peridiole_cross_section.svg.png)