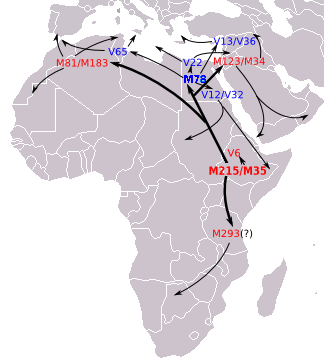
The ancient dispersals of the major E-M35 lineages. The map shows the supposed earliest movements of E-M215 lineages as described in the most recent articles.
In June 2015, Trombetta et al. reported a previously unappreciated large difference in the age between haplogroup E-M215 (38.6 kya; 95% CI 31.4–45.9 kya) and its sub-haplogroup E-M35 (25.0 kya; 95% CI 20.0–30.0 kya) and estimated its origin to be in Horn of Africa, where the node separating the E-V38 and E-M215 branches occurs about 47,500 years ago (95% CI: 41.3–56.8 ka).[2] E-M35 was dated by Batini in 2015 to between 15,400 and 20,500 years ago.[8]
All major sub-branches of E-M35 are thought to have originated in the same general area as the parent clade: in North Africa, the Horn of Africa, or nearby areas of the Near East. Some branches of E-M35 are assumed to have left Africa thousands of years ago, whereas others may have arrived from the Near East. For example, Underhill (2002) associates the spread of the haplogroup with the Neolithic Revolution, believing that the structure and regional pattern of E-M35 subclades potentially give "reagents with which to infer specific episodes of population histories associated with the Neolithic agricultural expansion". Battaglia et al. (2008) also estimate that E-M78 (called E1b1b1a1 in that paper) has been in Europe longer than 10,000 years. Accordingly, human remains excavated in a Spanish funeral cave dating from approximately 7,000 years ago were shown to be in this haplogroup.[9] Two more E-M78 have been found in the Neolithic Sopot and Lengyel cultures too.[10]
Concerning E-M35 in Europe within this scheme, Underhill & Kivisild (2007) have remarked that E-M215 seems to represent a late-Pleistocene migration from North Africa to Europe over the Sinai Peninsula in Egypt.[Note 2] While this proposal remains uncontested, it has more recently been proposed by Trombetta et al. (2011) that there is also evidence for additional migration of E-M215 carrying men directly from North Africa to southwestern Europe, via a maritime route (see below.)
Ancient DNA
According to Lazaridis et al. (2016), Natufian skeletal remains from the ancient Levant predominantly carried the Y-DNA haplogroup E. Of the five Natufian specimens analysed for paternal lineages, one belonged to CT, one to E-M35, one to E-M215, one to E-Z830.[11] Haplogroup E-M35 was also found the dominant marker among fossils from the ensuing Pre-Pottery Neolithic B culture, with the E-M78 and E-Z830 subclades observed in multiple PPNB specimens (~50%).[12]
Following Fregel et al. (2017), two Neolithic farmers from the North African site of Ifri n Ammar ou Moussa belonged to haplogroup E-L19. The majority of their genomes traced back to Natufian and PPN ancestry , indicating a recent intrusion into North Africa from the Levant. Their presence is likely linked to the spread of pastoral technologies from the Levant into North Africa.[13]
Loosdrecht et al. (2018) analysed genome-wide data from seven ancient Iberomaurusian individuals from the Grotte des Pigeons near Taforalt in eastern Morocco. The fossils were directly dated to between 15,100 and 13,900 calibrated years before present. The scientists found that five male specimens with sufficient nuclear DNA preservation all belonged to the E1b1b1a1 (M78) subclade, with one skeleton bearing the E1b1b1a1b1 parent lineage to E-V13.[14] Martiniano et al. (2022) later reassigned all the Taforalt samples to haplogroup E-M78 and none to E-L618, the predecessor to E-V13.[15]
Haplogroup E-M35 has been identified among both Amorite and Akkadian remains, found in the sites of Megiddo,[16] Ebla, and Alalakh.[17] It is also attested among later Semitic populations, such as the Phoenicians who settled throughout the Mediterranean region, where it appears to have been the most prevalent lineage.[18] Additionally, analyses of medieval Andalusian remains indicate that individuals examined for paternal DNA overwhelmingly belonged to the E-M81 subclade.[19][20][21]
In Egypt, haplogroup E-V1515 has been identified in the remains of an elite Egyptian individual from the site of Nuerat, dating to the Old Kingdom period,[22] while haplogroup E-V22 has been detected in an ancient Egyptian mummy excavated at the Abusir el-Meleq archaeological site in Middle Egypt, dated to between the late New Kingdom and the Roman era.[23]
Distribution
E-M215 and E-M35 are quite common among Afroasiatic speakers. The linguistic group and carriers of E-M35 lineage have a high probability to have arisen and dispersed together from the Afroasiatic Urheimat.[24] Amongst populations with an Afro-Asiatic speaking history, a significant proportion of Palestinians and Jewish male lineages are E-M35.[25] Haplogroup E-M35, which accounts for approximately 18%[3] to 20%[26][27] of 18% of Palestinians and Ashkenazi and 8.6%[28] to 30%[3] of Sephardi Y-chromosomes, appears to be one of the major founding lineages of the Palestinian and the Jewish population.[29][Note 3]
The following table only includes sample populations with more than 1% E-M215 men with all known subclades as of June 2015. It contains the E-V1515 clade defined by Trombetta et al. 2015, and all the E1b1b subclades distributed below the Sahara (E-V42, E-M293, E-V92, E-V6), which were identified as E-M35 basal clades in a former phylogeny.[2]
Population
N
Region
Language
Total E-M215
E-V2009
E-M78*
E-V1477
E-V1083*
E-V13
E-V22
E-V12*
E-V32
E-V259
E-V65
E-V257*
E-M81
E-M123*
E-M34
E-V1515*
E-V1486*
E-V2881*
E-V1792
E-V92
E-M293*
E-V3065
E-V42
E-V1785*
E-V6
E-V16
Northern Africa
Moroccan Arabs
55
Morocco
AA/Semitic
15.9
0.0
0.0
0.0
0.0
0.0
7.3
0.0
0.0
0.0
32.7
0.0
30.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Asni Berbers
54
Morocco
AA/Berber
85.2
0.0
0.0
0.0
0.0
0.0
3.7
0.0
0.0
0.0
0.0
1.9
79.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Bouhria Berbers
67
Morocco
AA/Berber
79.1
0.0
0.0
0.0
0.0
1.5
0.0
0.0
0.0
0.0
0.0
0.0
77.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Middle Atlas Berbers
69
Morocco
AA/Berber
81.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
10.1
0.0
71.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Marrakech Berbers
27
Morocco
AA/Berber
92.6
0.0
0.0
0.0
0.0
0.0
3.7
3.7
0.0
0.0
0.0
3.7
77.8
0.0
3.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Souss Berbers
34
Morocco
AA/Berber
79.4
2.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
76.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Ouarzazate Berbers
31
Morocco
AA/Berber
54.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
54.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Mozabite Berbers
67
Algeria
AA/Berber
89.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
1.5
0.0
86.6
0.0
1.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Tunisian Jews
10
Tunisia
AA/Semitic
20.0
0.0
0.0
10.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
10.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Libyan Arabs
10
Libya
AA/Semitic
20.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
20.0
0.0
30.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Libyan Jews
23
Libya
AA/Semitic
26.1
0.0
0.0
0.0
0.0
4.3
0.0
0.0
0.0
0.0
4.3
0.0
0.0
0.0
17.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Northern Egyptians
49
Egypt
AA/Semitic
20.9
0.0
0.0
0.0
0.0
2.0
16.3
4.1
2.0
0.0
0.0
0.0
4.1
4.1
10.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Egyptian Berbers from Siwa
93
Egypt
AA/Semitic
18.3
0.0
0.0
0.0
0.0
0.0
0.0
2.2
0.0
0.0
4.3
2.2
1.1
0.0
2.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
6.5
0.0
Egyptians from Baharia
41
Egypt
AA/Semitic
56.1
0.0
0.0
0.0
0.0
2.4
22.0
14.6
0.0
0.0
2.4
7.3
4.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
2.4
0.0
Egyptians from Gurna Oasis
34
Egypt
AA/Semitic
17.6
0.0
5.9
0.0
0.0
0.0
0.0
8.8
2.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southern Egyptians
47
Egypt
AA/Semitic
78.7
0.0
0.0
0.0
0.0
0.0
0.0
74.5
0.0
0.0
0.0
0.0
0.0
2.1
2.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Western/Central Africa
Mandenka
16
Senegal
NC/Mande
6.3
0.0
0.0
0.0
0.0
0.0
0.0
6.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Tuareg
22
Niger
AA/Berber
13.6
0.0
0.0
0.0
0.0
0.0
0.0
4.5
0.0
0.0
0.0
0.0
9.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Daba
29
Cameroon (North)
AA/Chadic
3.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Guidar
9
Cameroon (North)
AA/Chadic
11.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
11.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Mandara
82
Cameroon (North)
AA/Chadic
2.4
0.0
0.0
0.0
0.0
0.0
0.0
1.2
0.0
0.0
0.0
1.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Shuwa Arabs
5
Cameroon (North)
AA/Semitic
20.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
20.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Fulbe from Cameroon
76
Cameroon (North)
NC/Atlantic
1.3
1.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Moundang
21
Cameroon (North)
NC/Adamawa
4.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
4.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Eastern Africa
Tigre
5
Eritrea
AA/Semitic
100.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
60.0
0.0
0.0
0.0
0.0
0.0
0.0
20.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
20.0
0.0
Nara
15
Eritrea
NS/Sudanic
60.0
0.0
0.0
0.0
0.0
0.0
6.7
0.0
13.3
0.0
0.0
0.0
0.0
0.0
0.0
13.3
6.7
0.0
0.0
0.0
0.0
0.0
13.3
0.0
6.7
0.0
Cunama
20
Eritrea
NS/Cunama
65.0
0.0
0.0
0.0
0.0
0.0
5.0
0.0
20.0
0.0
0.0
0.0
0.0
0.0
5.0
0.0
0.0
5.0
0.0
0.0
0.0
0.0
15.0
10.0
5.0
0.0
Saho
94
Eritrea
AA/Cushitic
98.9
0.0
0.0
0.0
1.1
0.0
88.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
1.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
8.5
0.0
Tigrai
32
Eritrea/Ethiopia
AA/Semitic
71.9
0.0
0.0
0.0
0.0
0.0
3.1
3.1
21.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.1
3.1
0.0
0.0
0.0
0.0
0.0
31.3
6.3
0.0
Afar
25
Djibouti
AA/Cushitic
60.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
4.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
56.0
0.0
Somali
40
Djibouti
AA/Cushitic
25.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
25.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Ethiopian Jews
22
Ethiopia
AA/Cushitic
31.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
9.1
0.0
0.0
0.0
0.0
0.0
13.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
9.1
0.0
0.0
0.0
Amhara
82
Ethiopia
AA/Semitic
45.1
0.0
0.0
0.0
0.0
0.0
2.4
0.0
11.0
0.0
0.0
0.0
0.0
0.0
13.4
0.0
0.0
2.4
0.0
1.2
0.0
0.0
1.2
0.0
8.5
4.9
Oromo
62
Ethiopia
AA/Cushitic
53.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
22.6
0.0
0.0
0.0
0.0
0.0
4.8
0.0
0.0
17.7
0.0
0.0
1.6
0.0
3.2
0.0
1.6
1.6
Wolayta
12
Ethiopia
AA/Omotic
58.3
0.0
0.0
0.0
0.0
0.0
8.3
0.0
8.3
0.0
0.0
0.0
0.0
0.0
8.3
0.0
0.0
8.3
0.0
0.0
8.3
0.0
0.0
0.0
16.7
0.0
Somali
12
Ethiopia
AA/Cushitic
50.0
0.0
0.0
0.0
0.0
0.0
8.3
0.0
25.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
16.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Gurage
7
Ethiopia
AA/Semitic
42.9
0.0
0.0
0.0
0.0
0.0
28.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
14.3
0.0
Somali
5
Somalia
AA/Cushitic
100.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
80.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
20.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Turkana
6
Kenya
NS/Sudanic
50.0
0.0
0.0
0.0
0.0
0.0
33.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
16.7
0.0
0.0
0.0
0.0
0.0
Borana
9
Kenya
AA/Cushitic
77.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
66.7
0.0
0.0
11.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Somali
6
Kenya
AA/Cushitic
100.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
66.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
16.7
0.0
0.0
0.0
0.0
0.0
0.0
16.7
0.0
Nilotic Western Kenya
11
Kenya
NS/Sudanic
45.5
0.0
0.0
0.0
0.0
0.0
9.1
9.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
18.2
0.0
0.0
0.0
9.1
0.0
Luhya
51
Kenya
NC/Bantu
9.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
5.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.9
0.0
0.0
0.0
0.0
0.0
Other Bantu
17
Kenya
NC/Bantu
11.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
11.8
0.0
0.0
0.0
0.0
0.0
Kikuyu
9
Kenya
NC/Bantu
11.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
11.1
0.0
0.0
0.0
0.0
0.0
Maasai
45
Kenya
NS/Sudanic
37.8
0.0
0.0
0.0
0.0
0.0
6.7
0.0
6.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
24.4
0.0
0.0
0.0
0.0
0.0
Tutsi
9
Burundi
NC/Bantu
22.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
22.2
0.0
0.0
0.0
0.0
0.0
Southern Africa
!Kung
64
Angola
KS
10.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
9.4
1.6
0.0
0.0
0.0
0.0
Khwe
26
Namibia
KS
30.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
30.8
0.0
0.0
0.0
0.0
0.0
Bantu
8
South Africa
NC/Bantu
12.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
12.5
0.0
0.0
0.0
0.0
0.0
Europe
Northern Portuguese
50
Portugal
IE
10.0
0.0
0.0
0.0
0.0
4.0
0.0
0.0
0.0
0.0
0.0
0.0
4.0
0.0
0.0
2.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southern Portuguese
49
Portugal
IE
16.3
0.0
0.0
0.0
0.0
4.1
0.0
0.0
0.0
0.0
0.0
0.0
12.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Pasiegos from Cantabria
56
Spain
IE
42.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
1.8
41.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Asturians
90
Spain
IE
12.2
0.0
0.0
0.0
0.0
5.6
4.4
0.0
0.0
0.0
0.0
0.0
2.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southern Spaniards
62
Spain
IE
6.5
0.0
0.0
0.0
0.0
0.0
3.2
0.0
0.0
0.0
0.0
1.6
1.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Spanish Basques
55
Spain
Basque
3.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
French
85
France
IE
8.2
0.0
0.0
0.0
0.0
3.5
0.0
1.2
0.0
0.0
0.0
0.0
3.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
French Basques
16
France
Basque
6.3
0.0
0.0
0.0
0.0
0.0
0.0
6.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Corsicans
140
France
IE
6.4
0.0
0.0
0.0
0.0
4.3
0.0
0.0
0.0
0.0
0.0
0.7
0.0
0.0
1.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Danish
35
Denmark
IE
2.9
0.0
0.0
0.0
0.0
2.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Germans
77
Germany
IE
3.9
0.0
0.0
0.0
0.0
3.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Northern Italians
80
Italy
IE
11.3
0.0
0.0
0.0
0.0
6.3
2.5
0.0
0.0
0.0
0.0
0.0
1.3
0.0
1.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Central Italians
356
Italy
IE
12.9
0.0
0.0
0.0
0.0
5.3
2.0
0.3
0.0
0.0
0.3
0.3
0.8
0.0
3.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southern Italians
141
Italy
IE
15.6
0.7
0.0
0.0
0.0
8.5
1.4
0.7
0.0
0.0
0.0
0.0
1.4
0.0
2.8
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Sicilians
153
Italy
IE
20.3
0.0
0.0
0.0
0.0
7.2
4.6
0.7
0.0
0.0
0.7
0.0
0.7
0.0
6.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Sardinians
374
Italy
IE
8.3
0.8
0.0
0.0
0.3
1.1
0.8
0.3
0.0
0.0
1.1
0.3
0.3
0.0
3.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Polish
40
Poland
IE
2.5
0.0
0.0
0.0
0.0
2.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Slovenians
104
Slovenia
IE
2.9
0.0
0.0
0.0
0.0
2.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Estonians
74
Estonia
U
5.4
0.0
0.0
0.0
0.0
4.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Hungarians
106
Hungary
U
10.4
0.0
0.0
0.0
0.0
9.4
0.0
0.0
0.0
0.0
0.0
0.0
0.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Romanians
30
Romania
IE
26.7
0.0
0.0
0.0
0.0
26.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Macedonians
99
Macedonia
IE
18.2
0.0
0.0
0.0
0.0
18.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Continental Greeks
32
Greece
IE
28.1
0.0
0.0
0.0
0.0
25.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Bulgarians
112
Bulgaria
IE
22.3
0.0
0.0
0.0
0.0
21.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Sephardic Bulgarians
20
Bulgaria
IE
5.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
5.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Albanians
21
Albania
IE
33.3
0.0
0.0
0.0
0.0
33.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Near East
Sephardic Turkish
19
Turkey
A
10.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
5.3
0.0
5.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Istanbul Turkish
35
Turkey
A
17.1
0.0
0.0
0.0
0.0
2.9
5.7
0.0
0.0
0.0
0.0
0.0
5.7
0.0
2.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southwestern Turkish
40
Turkey
A
7.5
0.0
0.0
0.0
0.0
2.5
0.0
0.0
0.0
0.0
0.0
0.0
2.5
0.0
2.5
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Northeastern Turkish
41
Turkey
A
2.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
2.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Central Anatolian
61
Turkey
A
9.8
0.0
0.0
0.0
0.0
4.9
0.0
1.6
0.0
0.0
0.0
0.0
0.0
0.0
3.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Southeastern Turkish
24
Turkey
A
8.3
0.0
0.0
0.0
0.0
4.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
4.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Erzurum Turkish
25
Turkey
A
12.0
0.0
0.0
0.0
0.0
0.0
0.0
4.0
0.0
0.0
0.0
0.0
0.0
0.0
8.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Turkish Cypriots
46
Turkey
A
23.9
0.0
0.0
0.0
0.0
10.9
2.2
0.0
0.0
0.0
0.0
0.0
8.7
0.0
2.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Bedouins
28
Israel
AA/Semitic
14.3
0.0
0.0
0.0
0.0
0.0
3.6
0.0
0.0
0.0
0.0
0.0
3.6
0.0
7.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Druze Arabs
28
Israel
AA/Semitic
14.3
0.0
0.0
0.0
0.0
10.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.6
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Palestinians
29
Israel
AA/Semitic
13.8
0.0
0.0
0.0
0.0
3.4
6.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
3.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Emiratis
41
United Arab Emirates
AA/Semitic
7.3
0.0
0.0
0.0
0.0
0.0
2.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
4.9
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Omanites
13
Oman
AA/Semitic
15.4
0.0
0.0
0.0
0.0
0.0
7.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
7.7
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
Yemenites
94
Yemen
AA/Semitic
14.9
0.0
0.0
0.0
0.0
0.0
0.0
2.1
3.2
0.0
0.0
0.0
1.1
0.0
7.4
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
1.1
Exceptional cases of men who are M215 positive but M35 negative ("E-M215*") have been discovered so far in two Amharas of Ethiopia and one Yemeni.[4][30] At least some of these men, perhaps all, are known since early 2011 to be in a rare sibling clade to E-M35, known as E-V16 or E-M281.[31] The discovery of M281 was announced by Semino et al. (2002), who found it in two Ethiopian Oromo. Trombetta et al. (2011) found 5 more Ethiopian individuals and an equivalent SNP to M281, V16. It was in the 2011 paper that the family tree position (M215+/M35-) was discovered as described above. The E-M215 derivative, E-M35 is defined by the M35 SNP. 1 Turkmen individual from Jawzjan with a subclade defining mutation is referred to as E-M35*.[32] As of June 2015, there is an increasingly complex tree structure which divides most men in E-M35 into two branches: E-V68 and E-Z827.
The most frequently described subclades are E-M78, a part of E-V68, and E-M81, which is a branch of E-Z827. These two subclades represent the largest proportion of the modern E-M215 population. E-M78 is found over most of the range where E-M215 is found excluding Southern Africa. E-M81 is found mainly in North Africa. E-M123 is less common but widely scattered, with significant populations in specific parts of the Horn of Africa, the Levant, Arabia, Iberia, and Anatolia. A new clade (E-V1515) was defined by Trombetta et al. 2015, which originated about 12 kya (95% CI 8.6–16.4) in eastern Africa where it is currently mainly distributed. This clade includes the E-V42, E-M293, E-V92 and E-V6 subclades, which were identified as E-M35 basal clades in a previous phylogeny.[2]
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257. Both contain a lineage which has been frequently observed in Africa (E-M78 and E-M81, respectively) and a group of undifferentiated chromosomes that are mostly found in southern Europe. An expansion of E-M35 carriers, possibly from the Middle East as proposed by other authors, and split into two branches separated by the geographic barrier of the Mediterranean Sea, would explain this geographic pattern. However, the absence of E-V68* and E-V257* in the Middle East makes a maritime spread between northern Africa and southern Europe a more plausible hypothesis.
E-V68, is dominated by its longer-known subclade E-M78. Three "E-V68*" individuals who are in E-V68 but not E-M78 have been reported in Sardinia, by Trombetta et al. (2011), when announcing the discovery of V68. The authors noted that because E-V68* was not found in the Middle Eastern samples, this appears to be evidence of maritime migration from Africa to southwestern Europe. E-M78 is a commonly occurring subclade, widely distributed in North Africa, the Horn of Africa, West Asia, (the Middle East and Near East) "up to Southern Asia",[5] and all of Europe.[33] The European distribution has a frequency peak centered in parts of the Balkans (up to almost 50% in some areas)[3][34] and Sicily, and declining frequencies evident toward western, central, and northeastern Europe.
Based on genetic STR variance data, Cruciani et al. (2007) suggests that E-M78 originated in the region of Egypt and Libya.[Note 4] about 18,600 years ago (17,300 – 20,000 years ago).[Note 5]Battaglia et al. (2008) describe Egypt as "a hub for the distribution of the various geographically localized M78-related subclades" and, based on archaeological data, they propose that the point of origin of E-M78 (as opposed to later dispersal from Egypt) may have been in a refugium which "existed on the border of present-day Sudan and Egypt, near Lake Nubia, until the onset of a humid phase around 8500 BC. The northward-moving rainfall belts during this period could have also spurred a rapid migration of Mesolithic foragers northwards in Africa, the Levant and ultimately onward to Asia Minor and Europe, where they each eventually differentiated into their regionally distinctive branches". Towards the south, Hassan et al. (2008) also explain evidence that some subclades of E-M78, specifically E-V12 and E-V22, "might have been brought to Sudan from North Africa after the progressive desertification of the Sahara around 6,000–8,000 years ago". And similarly, Cruciani et al. (2007) propose that E-M78 in Ethiopia, Somalia and surrounding areas, back-migrated to this region from the direction of Egypt after acquiring the E-M78 mutation.
Recently, E-M78 was dated by Trombetta et al. 2015. between 20,300 and 14,800 years ago.[2]
Subclades of E-M78
Listed here are the main subclades of M78 as of June 2015. Within the E-M78 subclade, Trombetta et al. 2015 allocated most of the former E-M78* chromosomes to three new distinct branches: E-V1083*, E-V1477 and E-V259. The first is a paragroup sister to clades E-V22 and E-V13. The mutation V1477 defines a new basal branch that has been observed only in one northern African sample. Finally, a sister clade of E-V12 defined by V264 includes E-V65 and V259, a new lineage distributed in central Africa.[2][33]
E-M78 (E1b1b1a1) North Africa, Horn of Africa, West Asia, Europe (formerly "E1b1b1a").
E-M78* Found in Morocco, southern Portugal, southern Spain and Iran (Tehran and Semnan provinces).
E-V1477 Found in Tunisian Jews.
E-V1083
E-V1083* Found only in Eritrea (1.1%) and Sardinia (0.3%).
E-V13 This is the most common subclade of E-M215 found in Europe. It is especially common in the Balkans.
E-V22. Concentrated in Northeast Africa and the Near East. Peaks among the Saho.
E-V1129
E-V12. Found in Egypt, Sudan, and Chad other places. Has an important subclade
E-V12* Most common lineage among Southern Egyptians (74.5%).
E-V32. Very common among Somalis, Tigre and Oromos.
E-V264
E-V259 Found in North Cameroon.
E-V65 Associated with North Africa, but also found in Sicily and also found in continental Italy.
E-M521 Not mentioned by Trombetta et al.2015. Found in two individuals in Greece by Battaglia et al. (2008) and in one individual from the Eastern Alpine region of Italy by Coia et al. (2013)
E-V257* individuals in their samples who were E-V257, but not E-M81. A Borana from Kenya, a Marrakesh Berber, a Corsican, a Sardinian, a southern Spaniard and a Cantabrian. As mentioned above, Trombetta et al. (2011) propose that the absence of E-V257* in the Middle East (Yfull found a young one in Iranian Azerbaijan[35] and a different young one in Armenia[36]) makes a maritime movement from northern Africa to southern Europe the most plausible hypothesis so far to explain its distribution. Yfull lists 24 individuals, all of whom belong to a single branch that is 30% younger than their common ancestor with M81.[37]
E-M81
E-M81 is the most common subclade of haplogroup E-L19/V257. It is concentrated in the Maghreb, and is dominated by its E-M183 subclade. E-M183 is believed to have originated in the Northwest Africa and has an estimated age of 4700 ybp.[38]
This haplogroup reaches a mean frequency of 61% in the Maghreb and 51% in North Africa, decreasing in frequency from approximately 80% to 100% in Berber populations,[39][40] including Saharawis, to approximately 29% to the east of this range in Egypt.[41][42][43][44] Because of its young age and prevalence among these groups and also others such as Mozabite, Middle Atlas, Kabyle and other Berber groups, it is sometimes referred to as a genetic "Berber marker". Pereira et al. (2010) report high levels amongst Tuareg in two Saharan populations – 77.8% near Gorom-Gorom, in Burkina Faso, and 81.8% from Gosi in Mali. There was a much lower frequency of 11.1% in the vicinity of Tanut in Niger. E-M81 is also quite common among Maghrebi Arabic-speaking groups. It is generally found at frequencies around 45% in coastal cities of Algeria and Tunisia (Jijel, Oran, Tizi Ouzou, Algiers, Tunis, Sousse).[41][45]
In this key area from Egypt to the Atlantic Ocean, Solé-Morata et al. (2017) report a M183-SM001 pattern of decreasing microsatellite haplotype variation (implying greater lineage age in the former areas) from the Reguibat tribe in Oran and they found M183* (not SM001) in Iberia, Libya and Morocco. Arredi et al. (2004) however showed microsatellite variation decrease from East to West, accompanied by a substantial increasing frequency. At the eastern extreme of this core range, Kujanova et al. (2009) found M81 in 28.6% (10 out of 35 men) in el-Hayez in the Libyan Desert in Egypt.
Arredi et al. (2004) believe the pattern of distribution and variance to be consistent with the hypothesis of a "demic diffusion" from the East. There is no autochthonous presence of E-M81 in the Near East (there is one in Lebanon[46]), indicating that M81 most likely emerged from its parent clade M35 either in North Africa, or possibly as far south as the Horn of Africa.[47]
In Europe, E-M81 is widespread but rare, in the Iberian Peninsula shows an average frequency of 4% (45/1140) in the Iberian Peninsula with frequencies reaching 3.5% in Galicia, 4% in Western Andalusia and Northwest Castile. However this study includes 153 individuals from Majorca, Minorca and Ibiza islands as well as 24 individuals from Gascony which are not in the Iberian Peninsula. Without these 177 individuals, average for Iberian Peninsula is 4.1% (40/963),[48] it is found at comparable levels to E-M78, with an average frequency of around 5%. Its frequencies are higher in the western half of the peninsula with frequencies reaching 8% in Extremadura and southern Portugal, 4% to 5% in Galicia, 5% in western Andalusia and 4% in northwest Castile and 9% to 17% in Cantabria.[28][49][50][51][52] The highest frequencies of this clade found so far in Europe were observed in the Valles Pasiegos from Cantabria, ranging from 5.5% (8/45)[52] to 41% (23/56).[4] An average frequency of 8.28% (54/652) has also been reported in the Spanish Canary Islands with frequencies over 10% in the three largest islands of Tenerife (10.68%), Gran Canaria (11.54%) and Fuerteventura (13.33%).[53]
E-M81 is also found in France,[4] 2.70% (15/555) overall with frequencies surpassing 5% in Auvergne (5/89) and Île-de-France (5/91),[54][55] 0,7% to 5,8% in Sardinia,[56][57] approximately 2.12% overall in Sicily (but up to 7.14% in Piazza Armerina),[58] and in very much lower frequency near Lucera (1.7%), in continental Italy,[51] possibly due to ancient migrations during the Islamic, Roman, and Carthaginian empires. In a 2014 study by Stefania Sarno et al. with 326 samples from Cosenza, Reggio Calabria, Lecce and five Sicilian provinces, E-M81 shows an average frequency of 1.53%, but the typical Maghrebin core haplotype 13-14-30-24-9-11-13 has been found in only two out of the five E-M81 individuals. These results, along with the negligible contribution from North-African populations revealed by the admixture-like plot analysis, suggest only a marginal impact of trans-Mediterranean gene flows on the current SSI genetic pool.[58][59]
In smaller numbers, E-M81 men can be found in areas in contact with North Africa, both around the Sahara, in places like Sudan, and around the Mediterranean in places like Lebanon, Turkey, and amongst Sephardi Jews.
There are two recognized subclades of E-M81, although one is much more important than the other.[which?]
The E-M81 subclade has been found in ancient Guanche (Bimbapes) fossils excavated in Punta Azul, El Hierro, Canary Islands, which are dated to the 10th century (~44%).[62]
E-M183 is extremely dominant (more than 99%[63]) within E-M81. Karafet et al. (2008) first described it as a subclade of E-M81. The known subclades of E-M183 include:
This is a recently discovered subclade which has not yet been included in most haplogroup trees, E-Z830 includes the confirmed subclades of E-M123, E-V1515 (E-M293, E-V42, E-V6, E-V92), and E-Z830*, and is a sibling clade to E-L19. Currently,[64] the E-M35 phylogeny project] recognizes four distinct clusters of Z830* carriers, two of which are exclusively Jewish in origin. The remaining two are significantly smaller, and include scattered individuals in Germany, Spain, Latin America, Egypt, and Ethiopia.[65][66][67][68]
E-PF2431
E-PF2431 is a minor subclade of E-L19 mostly found in Europe, with smaller but notable frequencies in the Middle East/North Africa and sub-Saharan Africa. This haplogroup has been identified among Phoenicians from Motya,[69] ancient Romans from Pompeii,[70] and Medieval Arabs.[71]
E-Z830 (E1b1b1b2)
This is a recently discovered subclade which has not yet been included in most haplogroup trees, E-Z830 includes the confirmed subclades of E-M123, E-V1515 (E-M293, E-V42, E-V6, E-V92), and E-Z830*, and is a sibling clade to E-L19. Currently,[72] the E-M35 phylogeny project] recognizes four distinct clusters of Z830* carriers, two of which are exclusively Jewish in origin. The remaining two are significantly smaller, and include scattered individuals in Germany, Spain, Latin America, Egypt, and Ethiopia.[73][74][75][76]
E-M123 is mostly known for its major subclade E-M34, which dominates this clade.[Note 7]
E-V1515
A new clade (E-V1515) was defined by Trombetta et al. 2015, which originated about 12 kya (95% CI 8.6–16.4) in eastern Africa where it is currently mainly distributed. This clade includes all the sub-Saharan haplogroups (E-V42, E-M293, E-V92, E-V6) reported as E-M35 basal clades in a previous phylogeny.[2]
E-M293
E-M293 is a subclade of E-V1515. It was first identified by ISOGG as the second clade within E-Z830. It was discovered before E-Z830, being announced in Henn et al. (2008), which associated it with the spread of pastoralism from East Africa into Southern Africa. So far high levels have been found in specific ethnic groups in Tanzania and Southern Africa. Highest were the Datooga (43%), Khwe (Kxoe) (31%), Burunge (28%), and Sandawe peoples (24%). Henn (2008) in their study also found two Bantu-speaking Kenyan males with the M293 mutation.[6]
Other E-M215 subclades are rare in Southern Africa. The authors state "Without information about M293 in the Maasai, Hema, and other populations in Kenya, Sudan, and Ethiopia, we cannot pinpoint the precise geographic source of M293 with greater confidence. However, the available evidence points to present-day Tanzania as an early and important geographic locus of M293 evolution.". They also say that "M293 is only found in sub-Saharan Africa, indicating a separate phylogenetic history for M35.1 * (former) samples further north".
E-P72 appears in Karafet (2008). Trombetta et al. (2011) announced that this is a subclade of E-M293.
E-V42
Trombetta et al. (2011) announced the discovery of E-V42 in two Beta Israel persons. It was suggested that it may be restricted to the region around Ethiopia. However, further testing by commercial DNA testing companies confirmed many positive results for this subclade in Saudi Arabia, Kuwait and one person in Portugal who has a root from Arabia.[77]
E-V6
The E-V6 subclade of E-V1515 is defined by V6. Cruciani et al. (2004) identified a significant presence of these lineages in Ethiopia and also some in the neighboring Somalis. Among the Ethiopian and Somali samples, the highest were 14.7% among the Amhara and 16.7% among the Wolayta.
To the south, Tishkoff et al. (2007) identified one V6+ man in a sample of 35 Datooga of Tanzania. And further to the north, Dugoujon et al. (2009) identified another 6 men in a sample of 93 from the Siwa Oasis, which is a Berber population
E-V92
Trombetta et al. (2011) announced the discovery of E-V92 in two Amharas. Like E-V6 and E-V42 it possibly only exists in the area of Ethiopia.
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
The following phylogenetic tree is based on the YCC 2008 tree and subsequent published research as summarized by ISOGG. It includes all known subclades as of June 2015 (Trombetta et al. 2015)[78][79][80]
↑Van Oven M, Van Geystelen A, Kayser M, Decorte R, Larmuseau HD (2014). "Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome". Human Mutation. 35 (2): 187–91. doi:10.1002/humu.22468. PMID24166809. S2CID23291764.
↑K-M2313*, which as yet has no phylogenetic name, has been documented in two living individuals, who have ethnic ties to India and South East Asia. In addition, K-Y28299, which appears to be a primary branch of K-M2313, has been found in three living individuals from India. See: Poznik op. cit.; YFull YTree v5.08, 2017, "K-M2335", and; PhyloTree, 2017, "Details of the Y-SNP markers included in the minimal Y tree" (Access date of these pages: 9 December 2017)
↑ Haplogroup M, as of 2017, is also known as K2b1b. (Previously the name Haplogroup M was assigned to K2b1d.)
↑ Haplogroup S, as of 2017, is also known as K2b1a. (Previously the name Haplogroup S was assigned to K2b1a4.)
Notes
↑Semino et al. (2004): "Both phylogeography and microsatellite variance suggest that E-P2 and its derivative, E-M35, probably originated in eastern Africa."
↑"Y chromosome data show a signal for a separate late-Pleistocene migration from Africa to Europe via Sinai as evidenced through the distribution of haplogroup E3b lineages, which is not manifested in mtDNA haplogroup distributions."Underhill & Kivisild (2007:547)
↑"Paragroup E-M35 * and haplogroup J-12f2a* fit the criteria for major AJ founding lineages because they are widespread both in AJ populations and in Near Eastern populations, and occur at much lower frequencies in European non-Jewish populations." Behar et al. (2004)
↑Cruciani et al. (2007) use the term Northeastern Africa to refer to Egypt and Libya, as shown in Table 1 of the study. Prior to Cruciani et al. (2007), Semino et al. (2004) East Africa as a possible place of origin of E-M78, based upon Ethiopian testing. This was because of the high frequency and diversity of E-M78 lineages in the region of Ethiopia. However, Cruciani et al. (2007) were able to study more data, including populations from North Africa who were not represented in the Semino et al. (2004) study, and found evidence that the E-M78 lineages which make up a significant proportion of some populations in that region, were relatively young branches (see E-V32 below). They therefore concluded that "Northeast Africa" was the likely place of origin of E-M78 based on "the peripheral geographic distribution of the most derived subhaplogroups with respect to northeastern Africa, as well as the results of quantitative analysis of UEP and microsatellite diversity". So according to Cruciani et al. (2007) E-M35, the parent clade of E-M78, originated in East Africa, subsequently spread to Northeast Africa, and then there was a "back migration" of E-M215 chromosomes that had acquired the E-M78 mutation. Cruciani et al. (2007) therefore note this as evidence for "a corridor for bidirectional migrations" between Northeast Africa (Egypt and Libya in their data) on the one hand and East Africa on the other. The authors believe there were "at least 2 episodes between 23.9–17.3 ky and 18.0–5.9 ky ago".
↑Cruciani et al. (2007) use two calculation methods for estimating the age of E-M78 which give very different results. For the main 18,600 years ago, the ASD method is used, while for a second "Ρ method", used as a check, gives 13.7kya with a Standard deviation of 2.3kya, but the difference between the two methods is only large for the age estimation of E-M78, not its subclades. The authors state that the big difference is "attributable to the relevant departure from a star-like structure because of repeated founder effects"
↑(6 out of 112), "The presence of chromosomes of North African origin (E3b1b-M81; Cruciani et al., 2004) can also be explained by a Portuguese but especially Italian-mediated influx, since this haplogroup reaches a frequency of 4.6% in Portugal and of 4.8% Italy, quite similar to the frequency found in Rio de Janeiro (4.4%) among European contributors." Silva et al. (2006)
↑Fregel, R.; Méndez, F. L.; Bokbot, Y.; Martín-Socas, D.; Camalich-Massieu, M. D.; Santana, J.; Morales, J.; Ávila-Arcos, M. C.; Underhill, P. A.; Shapiro, B.; Wojcik, G.; Rasmussen, M.; Soares AER; Kapp, J.; Sockell, A.; Rodríguez-Santos, F. J.; Mikdad, A.; Trujillo-Mederos, A.; Bustamante, C. D. (2018). "PNAS". Proceedings of the National Academy of Sciences of the United States of America. 115 (26): 6774–6779. doi:10.1073/pnas.1800851115. PMC6042094. PMID29895688.
↑Kujanová, Martina; Pereira, Luísa; Fernandes, Verónica; Pereira, Joana B.; Cerný, Viktor (October 2009). "Near eastern neolithic genetic input in a small oasis of the Egyptian Western Desert". American Journal of Physical Anthropology. 140 (2): 336–346. Bibcode:2009AJPA..140..336K. doi:10.1002/ajpa.21078. ISSN1096-8644. PMID19425100.
↑Ordóñez, A. C.; Fregel, R.; Trujillo-Mederos, A.; Hervella, M.; de-la-Rúa, C.; Arnay-de-la-Rosa, M. (2017). "Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)". Journal of Archaeological Science. 78: 20–28. Bibcode:2017JArSc..78...20O. doi:10.1016/j.jas.2016.11.004.
Adams, Susan M; Bosch, Elena; Balaresque, Patricia L.; Ballereau, Stéphane J.; Lee, Andrew C.; Arroyo, Eduardo; López-Parra, Ana M.; Aler, Mercedes; etal. (2008), "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula", The American Journal of Human Genetics, 83 (6): 725–36, doi:10.1016/j.ajhg.2008.11.007, PMC2668061, PMID19061982
Alvarez; Santos, Cristina; Montiel, Rafael; Caeiro, Blazquez; Baali, Abdellatif; Dugoujon, Jean-Michel; Aluja, Maria Pilar (2009), "Y-chromosome variation in South Iberia: Insights into the North African contribution", American Journal of Human Biology, 21 (3): 407–409, doi:10.1002/ajhb.20888, PMID19213004, S2CID7041905
Badro, Danielle A.; Douaihy, Bouchra; Haber, Marc; Youhanna, Sonia C.; Salloum, Angélique; Ghassibe-Sabbagh, Michella; Johnsrud, Brian; Khazen, Georges; Matisoo-Smith, Elizabeth; Soria-Hernanz, David F.; Wells, R. Spencer; Tyler-Smith, Chris; Platt, Daniel E.; Zalloua, Pierre A. (2013), "Y-Chromosome and mtDNA Genetics Reveal Significant Contrasts in Affinities of Modern Middle Eastern Populations with European and African Populations", PLOS ONE, 8 (1: e54616) e54616, Bibcode:2013PLoSO...854616B, doi:10.1371/journal.pone.0054616, PMC3559847, PMID23382925
Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri; etal. (2008), "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe", European Journal of Human Genetics, 17 (6): 820–830, doi:10.1038/ejhg.2008.249, PMC2947100, PMID19107149
Bosch, Elena; Calafell, Francesc; Comas, David; Oefner, Peter J.; Underhill, Peter A.; Bertranpetit, Jaume (2001), "High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between north-western Africa and the Iberian Peninsula", Am J Hum Genet, 68 (4): 1019–1029, doi:10.1086/319521, PMC1275654, PMID11254456
Cadenas; Zhivotovsky, Lev A; Cavalli-Sforza, Luca L; Underhill, Peter A; Herrera, Rene J (2007), "Y-chromosome diversity characterizes the Gulf of Oman", European Journal of Human Genetics, 16 (3): 1–13, doi:10.1038/sj.ejhg.5201934, PMID17928816
Caratti; Gino, S.; Torre, C.; Robino, C. (2009), "Subtyping of Y-chromosomal haplogroup E-M78 (E1b1b1a) by SNP assay and its forensic application", International Journal of Legal Medicine, 123 (4): 357–360, doi:10.1007/s00414-009-0350-y, PMID19430804, S2CID5657112
Capelli, Cristian; Onofri, Valerio; Brisighelli, Francesca; Boschi, Ilaria; Scarnicci, Francesca; Masullo, Mara; Ferri, Gianmarco; Tofanelli, Sergio; etal. (2009), "Moors and Saracens in Europe: estimating the medieval North African male legacy in southern Europe", European Journal of Human Genetics, 17 (6): 848–852, doi:10.1038/ejhg.2008.258, PMC2947089, PMID19156170
Contu, Daniela; Morelli, Daniela; Santoni, Federico; Foster, Jamie W.; Francalacci, Paolo; Cucca, Francesco (2008), "Y-Chromosome Based Evidence for Pre-Neolithic Origin of the Genetically Homogeneous but Diverse Sardinian Population: Inference for Association Scans", PLOS ONE, 3 (1) e1430, Bibcode:2008PLoSO...3.1430C, doi:10.1371/journal.pone.0001430, PMC2174525, PMID18183308
Cruciani, Fulvio; Santolamazza, Piero; Shen, Peidong; MacAulay, Vincent; Moral, Pedro; Olckers, Antonel; Modiano, David; Holmes, Susan (2002), "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes", American Journal of Human Genetics, 70 (5): 1197–1214, Bibcode:2002AmJHG..70.1197C, doi:10.1086/340257, PMC447595, PMID11910562
Cruciani; La Fratta; Torroni; Underhill; Scozzari (2006), "Molecular Dissection of the Y Chromosome Haplogroup E-M78 (E3b1a): A Posteriori Evaluation of a Microsatellite-Network-Based Approach Through Six New Biallelic Markers", Human Mutation, 27 (8): 831–2, doi:10.1002/humu.9445, PMID16835895, S2CID26886757
Di Gaetano; Cerutti, Francesca; Crobu, Carlo; Robino (2009), "Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome", European Journal of Human Genetics, 17 (1): 91–99, doi:10.1038/ejhg.2008.120, PMC2985948, PMID18685561
El-Sibai, Mirvat; Platt, Daniel E.; Haber, Marc; Xue, Yali; Youhanna, Sonia C.; Wells, R. Spencer; Izaabel, Hassan; Sanyoura, May F.; etal. (2009), "Geographical Structure of the Y-chromosomal Genetic Landscape of the Levant: A coastal-inland contrast", Annals of Human Genetics, 73 (6): 568–581, doi:10.1111/j.1469-1809.2009.00538.x, PMC3312577, PMID19686289
Firasat; Khaliq, Shagufta; Mohyuddin, Aisha; Papaioannou, Myrto; Tyler-Smith, Chris; Underhill, Peter A; Ayub, Qasim (2006), "Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan", European Journal of Human Genetics, 15 (1): 121–126, doi:10.1038/sj.ejhg.5201726, PMC2588664, PMID17047675
Flores, Carlos; Maca-Meyer, Nicole; González, Ana M; Oefner, Peter J; Shen, Peidong; Pérez, Jose A; Rojas, Antonio; Larruga, Jose M; Underhill, Peter A (2004), "Reduced genetic structure of the Iberian peninsula revealed by Y-chromosome analysis: implications for population demography", European Journal of Human Genetics, 12 (10): 855–863, doi:10.1038/sj.ejhg.5201225, PMID15280900, S2CID16765118
Flores; Maca-Meyer, Nicole; Larruga, Jose M.; Cabrera, Vicente M.; Karadsheh, Naif; Gonzalez, Ana M. (2005), "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan", J Hum Genet, 50 (9): 435–441, doi:10.1007/s10038-005-0274-4, PMID16142507
Francalacci, P.; Morelli, L.; Underhill, P.A.; Lillie, A.S.; Passarino, G.; Useli, A.; Madeddu, R.; Paoli, G.; etal. (2003), "Peopling of Three Mediterranean Islands (Corsica, Sardinia, and Sicily) Inferred by Y-Chromosome Biallelic Variability", American Journal of Physical Anthropology, 121 (3): 270–279, Bibcode:2003AJPA..121..270F, doi:10.1002/ajpa.10265, PMID12772214
Fregel, Rosa; Gomes, Verónica; Gusmão, Leonor; González, Ana M; Cabrera, Vicente M; Amorim, António; Larruga, Jose M (2009), "Demographic history of Canary Islands male gene-pool: replacement of native lineages by European", BMC Evolutionary Biology, 9 (1): 181, Bibcode:2009BMCEE...9..181F, doi:10.1186/1471-2148-9-181, PMC2728732, PMID19650893
Jobling, M.A.; Tyler-Smith, C. (2000), "New uses for new haplotypes the human Y chromosome, disease and selection", Trends Genet., 16 (8): 356–362, doi:10.1016/S0168-9525(00)02057-6, PMID10904265
King; Underhill (2002), "Congruent distribution of Neolithic painted pottery and ceramic figurines with Y-chromosome lineages", Antiquity, 76 (293): 707–14, doi:10.1017/S0003598X00091158, S2CID160359661
Kujanova; Pereira; Fernandes; Pereira; Cerný (2009), "Near Eastern Neolithic Genetic Input in a Small Oasis of the Egyptian Western Desert", American Journal of Physical Anthropology, 140 (2): 336–346, Bibcode:2009AJPA..140..336K, doi:10.1002/ajpa.21078, PMID19425100
Lacan, Marie; Keyser, Christine; Ricaut, François-Xavier; Brucato, Nicolas; Tarrús, Josep; Bosch, Angel; Guilaine, Jean; Crubézy, Eric; Ludes, Bertrand (2011), "Ancient DNA suggests the leading role played by men in the Neolithic dissemination", PNAS, 108 (45): 18255–9, Bibcode:2011PNAS..10818255L, doi:10.1073/pnas.1113061108, PMC3215063, PMID22042855
Maca-Meyer N, Sánchez-Velasco P, Flores C, Larruga JM, González AM, Oterino A, Leyva-Cobián F, etal. (2003), "Y Chromosome and Mitochondrial DNA Characterization of Pasiegos, a Human Isolate from Cantabria (Spain)", Annals of Human Genetics, 67 (Pt 4): 329–339, CiteSeerX10.1.1.584.4253, doi:10.1046/j.1469-1809.2003.00045.x, PMID12914567, S2CID40355653.
Martinez, Laisel; Underhill, Peter A; Zhivotovsky, Lev A; Gayden, Tenzin; Moschonas, Nicholas K; Chow, Cheryl-Emiliane T; Conti, Simon; Mamolini, Elisabetta; Cavalli-Sforza, L Luca; Herrera, Rene (April 1, 2007), "Paleolithic Y-haplogroup heritage predominates in a Cretan highland plateau", European Journal of Human Genetics, 15 (4): 485–493, doi:10.1038/sj.ejhg.5201769, ISSN1018-4813, PMID17264870
Mendizabal, Isabel; Sandoval, Karla; Berniell-Lee, Gemma; Calafell, Francesc; Salas, Antonio; Martinez-Fuentes, Antonio; Comas, David (2008), "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba", BMC Evol. Biol., 8 (1) 213, Bibcode:2008BMCEE...8..213M, doi:10.1186/1471-2148-8-213, PMC2492877, PMID18644108
Paracchini; Pearce, CL; Kolonel, LN; Altshuler, D; Henderson, BE; Tyler-Smith, C (2003), "A Y chromosomal influence on prostate cancer risk: the multi-ethnic cohort study", J Med Genet, 40 (11): 815–819, doi:10.1136/jmg.40.11.815, PMC1735314, PMID14627670
Pelotti; Ceccardi, S; Lugaresi, F; Trane, R; Falconi, M; Bini, C; Willuweit, S; Roewer, L (2007), "Microgeographic genetic variation of Y chromosome in a population sample of Ravenna's area in the Emilia-Romagna region (North of Italy)", Forensic Science International: Genetics Supplement Series, 1 (1): 242–243, doi:10.1016/j.fsigss.2007.10.025
Ramos-Luisa, E.; Blanco-Verea, A.; Brión, M.; Van Huffel, V.; Carracedo, A.; Sánchez-Diz, P. (2009), "Phylogeography of French male lineages (unpublished data 23rd International ISFG Congress)", Forensic Science International, 2: 439–441, doi:10.1016/j.fsigss.2009.09.026, S2CID85134429[dead link]
Robino, C.; Crobu, F.; Gaetano, C.; Bekada, A.; Benhamamouch, S.; Cerutti, N.; Piazza, A.; Inturri, S.; Torre, C. (2008), "Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample", Journal International Journal of Legal Medicine, 122 (3): 251–5, doi:10.1007/s00414-007-0203-5, PMID17909833, S2CID11556974
Rosa, Alexandra; Ornelas, Carolina; Jobling, Mark A; Brehm, António; Villems, Richard (2007), "Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective", BMC Evolutionary Biology, 7 (1): 124, Bibcode:2007BMCEE...7..124R, doi:10.1186/1471-2148-7-124, PMC1976131, PMID17662131
Sanchez, Juan J; Hallenberg, Charlotte; Børsting, Claus; Hernandez, Alexis; Gorlin, RJ (2005), "High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali males", European Journal of Human Genetics, 13 (7): 856–866, doi:10.1038/sj.ejhg.5201390, PMID15756297 . Published online 9 March 2005
Semino, Ornella; Magri, Chiara; Benuzzi, Giorgia; Lin, Alice A.; Al-Zahery, Nadia; Battaglia, Vincenza; MacCioni, Liliana; Triantaphyllidis, Costas; etal. (2004), "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area", American Journal of Human Genetics, vol.74, no.5, pp.1023–1034, Bibcode:2004AmJHG..74.1023S, doi:10.1086/386295, PMC1181965, PMID15069642
Shlush; Behar, Doron M.; Yudkovsky, Guennady; Templeton, Alan; Hadid, Yarin; Basis, Fuad; Hammer, Michael; Itzkovitz, Shalev; Skorecki, Karl (2008), Gemmell, Neil John (ed.), "The Druze: A Population Genetic Refugium of the Near East", PLOS ONE, 3 (5) e2105, Bibcode:2008PLoSO...3.2105S, doi:10.1371/journal.pone.0002105, PMC2324201, PMID18461126
Solé-Morata, Neus; García-Fernández, Carla; Urasin, Vadim; Bekada, Asmahan; Fadhlaoui-Zid, Karima; Zalloua, Pierre; Comas, David; Calafell, Francesc (21 November 2017), "Whole Y-chromosome sequences reveal an extremely recent origin of the most common North African paternal lineage E-M183 (M81)", Scientific Reports, 7 (1): 15941, Bibcode:2017NatSR...715941S, doi:10.1038/s41598-017-16271-y, ISSN2045-2322, PMC5698413, PMID29162904
Trombetta, Beniamino; Cruciani, Fulvio; Sellitto, Daniele; Scozzari, Rosaria (2011), MacAulay, Vincent (ed.), "A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms", PLOS ONE, 6 (1) e16073, Bibcode:2011PLoSO...616073T, doi:10.1371/journal.pone.0016073, PMC3017091, PMID21253605
Underhill, Peter A.; Shen, Peidong; Lin, Alice A.; Jin, Li; Passarino, Giuseppe; Yang, Wei H.; Kauffman, Erin; Bonné-Tamir, Batsheva; etal. (2000), "Y chromosome sequence variation and the history of human populations", Nat Genet, vol.26, no.3, pp.358–361, doi:10.1038/81685, PMID11062480, S2CID12893406
Underhill; Passarino, G.; Lin, A. A.; Shen, P.; Mirazon Lahr, M.; Foley, R. A.; Oefner, P. J.; Cavalli-Sforza, L. L. (2001), "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations", Ann Hum Genet, vol.65, no.Pt 1, pp.43–62, doi:10.1046/j.1469-1809.2001.6510043.x, PMID11415522, S2CID9441236
Underhill (2002), "Inference of Neolithic Population Histories using Y-chromosome Haplotypes", in Bellwood, Peter; Renfrew, A. Colin (eds.), Examining the farming/language dispersal hypothesis, Cambridge: McDonald Institute for Archaeological Research, ISBN978-1-902937-20-5
Underhill, Peter A.; Kivisild, Toomas (2007), "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations", Annu. Rev. Genet., 41: 539–64, doi:10.1146/annurev.genet.41.110306.130407, PMID18076332
Wood; Stover; Ehret; Destro-Bisol (2005), "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes", European Journal of Human Genetics, 13 (7): 867–876, doi:10.1038/sj.ejhg.5201408, PMID15856073
Y Chromosome Consortium "YCC" (2002), "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups", Genome Research, vol.12, no.2, pp.339–348, doi:10.1101/gr.217602, PMC155271, PMID11827954
Zalloua, Pierre A.; Xue, Yali; Khalife, Jade; Makhoul, Nadine; Debiane, Labib; Platt, Daniel E.; Royyuru, Ajay K.; Herrera, Rene J.; etal. (2008), "Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events", American Journal of Human Genetics, 82 (4): 873–882, doi:10.1016/j.ajhg.2008.01.020, PMC2427286, PMID18374297
Zalloua, Pierre A.; Platt, Daniel E.; El Sibai, Mirvat; Khalife, Jade; Makhoul, Nadine; Haber, Marc; Xue, Yali; Izaabel, Hassan; etal. (2008), "Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean", The American Journal of Human Genetics, 83 (5): 633–642, doi:10.1016/j.ajhg.2008.10.012, PMC2668035, PMID18976729
Zerjal (1999), The use of Y-chromosomal DNA variation to investigate population history; in Papiha SS, Deka R, Chakraborty R (eds): Genomic diversity: applications in human population genetics, Kluwer Academic/Plenum Publishers, pp.91–101
Zhao; Khan, Faisal; Borkar, Minal; Herrera, Rene; Agrawal, Suraksha (2009), "Presence of three different paternal lineages among North Indians: A study of 560 Y chromosomes", Annals of Human Biology, 36 (1): 1–14, doi:10.1080/03014460802558522, PMC2755252, PMID19058044
Underhill, Peter A.; Shen, Peidong; Lin, Alice A.; Jin, Li; etal. (November 2000). "Y chromosome sequence variation and the history of human populations". Nature Genetics. 26 (3): 358–361. doi:10.1038/81685. PMID11062480. S2CID12893406.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.

{kind=link}