
Lambeosaurinae is an extinct group of crested hadrosaurid dinosaurs.
Lambeosaurinae is an extinct group of crested hadrosaurid dinosaurs.

Uncertainty surrounds the size of lambeosaurs from the European continent. Hadrosaurs found there, alongside other dinosaurs, have traditionally been considered representatives of the phenomenon of insular dwarfism, as the continent was then made up of many smaller islands. Many fossil remains from the continent are smaller than those of hadrosaurs found elsewhere in the world, with only isolated remains indicating individuals of adult size by the standards of their relatives in North America and Asia. It remains possible, however, that at least some cases instead represent misidentification of juvenile remains. [2] [3] The presence of genuine dwarfed taxa has been validated in some cases; [2] [4] adults of the genus Minqaria , for example, are thought to be around 3.5 metres (11 ft) in length. [5] Contrastingly, the genus Pararhabdodon has a projected adult size similar to those of hadrosaurs on other continents, and known remains of Adynomosaurus and hadrosaurs from the Basturs Poble bonebed are of this adult size themselves. [6] Why hadrosaurs of such variable sizes co-exist, despite being subject to the same environmental pressures, remains unclear. [4]
Lambeosaurines originated on the continent of Laurasia during the Late Cretaceous, being initially found throughout modern Europe and Asia. Around the Campanian stage, lambeosaurines of the tribe Corythosauria colonized the landmass of Laramidia (modern western North America) via Beringia and spread as far south as Mexico, radiating into a diverse array of a body plans, including famous taxa such as Parasaurolophus and Lambeosaurus . For unknown reasons, lambeosaurines largely disappeared from North America around the Campanian/Maastrichtian boundary (the last remaining confirmed American member being Hypacrosaurus ), but continued their dominance in Laurasia up to the end of the Cretaceous, with some members such as Ajnabia and Minqaria even colonizing northern Africa from Europe. However, fragmentary remains, including a partial humerus, resembling those of lambeosaurines have been reported from the late Maastrichtian-aged New Egypt Formation of New Jersey, USA. If these are lambeosaurine remains, these specimens are unique both for representing the only potential record of lambeosaurines from the landmass of Appalachia (suggesting that lambeosaurines had managed to migrate eastwards to Appalachia during the Maastrichtian, following the partial closure of the Western Interior Seaway), and potentially representing one of the latest records of the group from North America. [7]

Lambeosaurines have been traditionally split into the tribes or clades Parasaurolophini ( Parasaurolophus , Charonosaurus , others (?).) and Lambeosaurini ( Corythosaurus , Hypacrosaurus , Lambeosaurus , others.). [8] Corythosaurini (synonym of Lambeosaurini, see below) and Parasaurolophini as terms entered the formal literature in Evans and Reisz's 2007 redescription of Lambeosaurus magnicristatus. Corythosaurini was defined as all taxa more closely related to Corythosaurus casuarius than to Parasaurolophus walkeri, and Parasaurolophini as all those taxa closer to P. walkeri than to C. casuarius. In this study, Charonosaurus and Parasaurolophus are parasaurolophins, and Corythosaurus, Hypacrosaurus, Lambeosaurus, Nipponosaurus , and Olorotitan are corythosaurins. [9] However, later researchers pointed out that due to the rules of priority set forth by the ICZN, Any tribe containing Lambeosaurus is properly named Lambeosaurini, and that therefore the name "Corythosaurini" is a junior synonym, and the definition had Corythosaurus casuarius changed to Lambeosaurus lambei, and the same for Parasaurolophini. [10] In more recent years Tsintaosaurini ( Tsintaosaurus + Pararhabdodon ) and Aralosaurini ( Aralosaurus + Canardia ) have also emerged. [11]
The following cladogram was recovered in a 2022 phylogenetic analysis by Xing Hai, and colleagues. [12]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
A 2013 study describing the genus Canardia found it to form a grouping with Aralosaurus , therein named as Aralosaurini and defined as the most exclusive clade of hadrosaurs containing both Canardia and Aralosaurus. [13] Though one 2022 study recovered the tribe as monophyletic, [12] most modern analyses fail to recover a natural grouping between the two genera. [14] [15] [16] Daniel Madzia and colleagues registered the name under Phylocode in a 2021 study and redefined it as "the largest clade containing Aralosaurus tuberiferus and Canardia garonnensis but not Lambeosaurus lambei, Parasaurolophus walkeri, and Tsintaosaurus spinorhinus". [17]
Numerous members of the clade Lambeosaurinae are known from Europe, dating to the end of the Cretaceous period. [13] These various named taxa have traditionally been found to group into numerous different lambeosaur lineages, including Aralosaurini, Tsintaosaurini, Lambeosaurini, [13] [15] and Parasaurolophini. [18] However, a study by Nick Longrich and colleagues proposed European lambeosaurs to form a singular monophyletic group, therein named Arenysaurini, based upon a phylogenetic analysis incorporating data based on geographic origin. They phylogenetically defined the tribe as all hadrosaurids closer to Arenysaurus ardevoli than Tsintaosaurus spinorhinus , Parasaurolophus walkeri , or Lambeosaurus lambei . In addition to the various named genera, indeterminate remains from across the continent including the Basturs Poble bonebed were proposed to represent arenysaurs. [15]
The existence of a tsintaosaur clade of lambeosaurines was first recognized by palaeontologists Albert Prieto-Márquez and Johnathan R. Wagner, who in 2009 published a paper recognizing a phylogenetic relationship between Tsintaosaurus and Pararhabdodon based both on shared anatomical traits and a phylogenetic analysis. [19] A 2013 study by Prieto-Márquez corroborated the existence of this grouping, and coined the tribe Tsintaosaurini to refer to it. The type genus is Tsintaosaurus, and it was defined as the smallest clade containing Tsintaosaurus spinorhinus and Pararhabdodon isonensis. [13] Several studies since have corroborated the existence of the clade, [17] [20] [21] though some others have failed to recover it, instead finding the two genera in a polytomy of basal lambeosaurs. [17] [22] [14] A 2021 paper by Daniel Madzia and other ornithischian researchers, focused on a revising ornithischian nomenclature and converting existing group names into Phylocode-compliant clades, re-formalized the coining of Tsintaosaurini and revised its definition to be the most inclusive group including T. spinorhinus and P. isonensis, but not Aralosaurus tuberiferus , Lambeosaurus lambei , or Parasaurolophus walkeri . [17] Longrich and colleagues instead consider the two genera unrelated, with Pararhabdodon being part of Arenysaurini. [15]
The term Corythosaurini was first used by Brett-Surman in 1989, who characterized the taxon via reference to the premaxilary expansion into a hollow helmet-like cranial crest, as well as higher neural spines. [23] The clade was formally defined via phylogenetic analysis by Evans and Reisz in 2007, [24] and this was confirmed by multiple other analyses. [25] In 2011, Sullivan et al. observed that by the rules of priority set by the International Code of Zoological Nomenclature, the name of the tribe ought to be Lambeosaurini due to its containing the defining type genus (Lambeosaurus) of its superior taxon (Lambeosaurinae). [26] It is defined as all lambeosaurines closer to Lambeosaurus lambei than to Parasaurolophus walkeri, Tsintaosaurus spinorhinus, or Aralosaurus tuberiferus. [17]

Hadrosaurids, or duck-billed dinosaurs, are members of the ornithischian family Hadrosauridae. This group is known as the duck-billed dinosaurs for the flat duck-bill appearance of the bones in their snouts. The ornithopod family, which includes genera such as Edmontosaurus and Parasaurolophus, was a common group of herbivores during the Late Cretaceous Period. Hadrosaurids are descendants of the Late Jurassic/Early Cretaceous iguanodontian dinosaurs and had a similar body layout. Hadrosaurs were among the most dominant herbivores during the Late Cretaceous in Asia and North America, and during the close of the Cretaceous several lineages dispersed into Europe, Africa, and South America.

Corythosaurus is a genus of hadrosaurid "duck-billed" dinosaur from the Late Cretaceous period, about 77–75.7 million years ago, in what is now western North America. Its name is derived from the Greek word κόρυς, meaning "helmet", named and described in 1914 by Barnum Brown. Corythosaurus is now thought to be a lambeosaurine, thus related to Lambeosaurus, Nipponosaurus, Velafrons, Hypacrosaurus, and Olorotitan. Corythosaurus has an estimated length of 7.7–9 metres (25–30 ft) and has a skull, including the crest, that is 70.8 centimetres tall.

Parasaurolophus is a genus of hadrosaurid "duck-billed" dinosaur that lived in what is now western North America and possibly Asia during the Late Cretaceous period, about 76.9–73.5 million years ago. It was a large herbivore that could reach over 9 metres (30 ft) long and weigh over 5 metric tons, and were able to move as a biped and a quadruped. Three species are universally recognized: P. walkeri, P. tubicen, and the short-crested P. cyrtocristatus. Additionally, a fourth species, P. jiayinensis, has been proposed, although it is more commonly placed in the separate genus Charonosaurus. Remains are known from Alberta, New Mexico, and Utah, as well as possibly Heilongjiang if Charonosaurus is in fact part of the genus. The genus was first described in 1922 by William Parks from a skull and partial skeleton found in Alberta.

Lambeosaurus is a genus of hadrosaurid dinosaur that lived about 75 million years ago, in the Late Cretaceous period of North America. This bipedal/quadrupedal, herbivorous dinosaur is known for its distinctive hollow cranial crest, which in the best-known species resembled a mitten. Several possible species have been named, from Canada, the United States, and Mexico, but only the two Canadian species are currently recognized as valid.
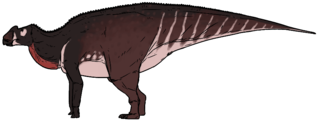
Aralosaurus was a genus of hadrosaurid dinosaur that lived during the Late Cretaceous in what is now Kazakhstan. It is known only by a posterior half of a skull and some post-cranial bones found in the Bostobe Formation in rocks dated from the Upper Santonian-Lower Campanian boundary, at about 83.6 Ma. Only one species is known, Aralosaurus tuberiferus, described by Anatoly Konstantinovich Rozhdestvensky in 1968. The genus name means Aral Sea lizard, because it was found to the northeast of the Aral Sea. The specific epithet tuberiferus means bearing a tuber because the posterior part of the nasal bone rises sharply in front of the orbits like an outgrowth. Aralosaurus was originally reconstituted with a nasal arch similar to that of North American Kritosaurus. For many years, Aralosaurus was thus placed in the clade of the Hadrosaurinae. This classification was invalidated in 2004, following the re-examination of the skull of the animal which allowed to identify in Aralosaurus many typical characters of Lambeosaurinae. In particular, this study revealed that Aralosaurus had a hollow bony structure located far in front of the orbits, which communicated with the respiratory tract. This structure being broken at its base, its shape and size remains undetermined. More recently, Aralosaurus has been identified as the most basal Lambeosaurinae, and placed with its close relative Canardia from the upper Maastrichtian of France in the new clade of Aralosaurini.

Amurosaurus is a genus of lambeosaurine hadrosaurid dinosaur found in the latest Cretaceous period of eastern Asia. Fossil bones of adults are rare, but an adult would most likely have been at least 6 metres (20 ft) long. According to Gregory S. Paul, it was about 8 metres (26 ft) long and weighed about 3,000 kilograms (6,600 lb).

Barsboldia is a genus of large hadrosaurid dinosaur from the early Maastrichtian Nemegt Formation of Ömnogöv', Mongolia. It is known from a partial vertebral column, partial pelvis, and some ribs.

Pararhabdodon is a genus of tsintaosaurin hadrosaurid dinosaur, from the Maastrichtian-age Upper Cretaceous Tremp Group of Spain. The first remains were discovered from the Sant Romà d’Abella fossil locality and assigned to the genus Rhabdodon, and later named as the distinct species Pararhabdodon isonensis in 1993. Known material includes assorted postcranial remains, mostly vertebrae, as well as maxillae from the skull. Specimens from other sites, including remains from France, a maxilla previously considered the distinct taxon Koutalisaurus kohlerorum, an additional maxilla from another locality, the material assigned to the genera Blasisaurus and Arenysaurus, and the extensive Basturs Poble bonebed have been considered at different times to belong to the species, but all of these assignments have more recently been questioned. It was one of the last non-avian dinosaurs known from the fossil record that went extinct during the Cretaceous-Paleogene extinction event.

Nipponosaurus is a lambeosaurine hadrosaur from sediments of the Yezo Group, in Sinegorsk on the island of Sakhalin, which was part of Japan at the time of the species' classification. The type and only species is N. sachalinensis, known only from a single juvenile specimen discovered in 1934 and named in 1936, by Takumi Nagao, with further material of the same individual found in 1937. Since then, the taxon has been largely ignored, and its validity has been doubted, with synonymy with other Asian hadrosaurs or status as a nomen dubium being suggested. Redescriptions from 2004 and 2017, however, have supported recognition as a distinct species. Dating the only specimen has been difficult, but based on associated mollusc taxa, the species likely lived sometime in the upper Santonian or lower Campanian, around 80 million years ago.

Prosaurolophus is a genus of hadrosaurid dinosaur from the Late Cretaceous of North America. It is known from the remains of at least 25 individuals belonging to two species, including skulls and skeletons, but it remains obscure. Its fossils have been found in the late Campanian-age Upper Cretaceous Dinosaur Park Formation in Alberta, and the roughly contemporaneous Two Medicine Formation in Montana, dating to around 75.5-74.0 million years ago. Its most recognizable feature is a small solid crest formed by the nasal bones, sticking up in front of the eyes.
Saurolophinae is a subfamily of hadrosaurid dinosaurs. It has since the mid-20th century generally been called the Hadrosaurinae, a group of largely non-crested hadrosaurs related to the crested sub-family Lambeosaurinae. However, the name Hadrosaurinae is based on the genus Hadrosaurus which was found in more recent studies to be more primitive than either lambeosaurines or other traditional "hadrosaurines", like Edmontosaurus and Saurolophus. As a result of this, the name Hadrosaurinae was dropped or restricted to Hadrosaurus alone, and the subfamily comprising the traditional "hadrosaurines" was renamed the Saurolophinae. Recent phylogenetic work by Hai Xing indicates that Hadrosaurus is placed within the monophyletic group containing all non-lambeosaurine hadrosaurids. Under this view, the traditional Hadrosaurinae is resurrected, with the Hadrosauridae being divided into two clades: Hadrosaurinae and Lambeosaurinae.

Koutalisaurus is a potentially dubious genus of extinct hadrosaurid dinosaur from the Arenysaurini. It is based on a mostly complete dentary from the Maastrichtian-age Upper Cretaceous Tremp Formation near the town of Abella de la Conca, Lleida, Spain.

Blasisaurus is a genus of lambeosaurine hadrosaurid dinosaur from the Late Cretaceous. It is known from a partial skull and skeleton found in late Maastrichtian-age rocks of Spain. The type species is Blasisaurus canudoi, described in 2010 by Penélope Cruzado-Caballero, Xabier Pereda-Suberbiola and José Ignacio Ruiz-Omeñaca, a group of researchers from Spain.

Magnapaulia is a genus of herbivorous lambeosaurine hadrosaurid dinosaurs known from the Latest Cretaceous Baja California, of northwestern Mexico. It contains a single species, Magnapaulia laticaudus. Magnapaulia was first described in 1981 as a possible species of Lambeosaurus by William J. Morris, and was given its own genus in 2012 by Prieto-Márquez and colleagues.

Latirhinus is an extinct genus of lambeosaurine hadrosaurid dinosaur from the Late Cretaceous of Mexico. The type species, Latirhinus uitstlani, was named in 2012 on the basis of a partial skeleton from the Campanian-age Cerro del Pueblo Formation. The specific name uitstlani means "southern" in the Náhuatl language of Mexico, a reference to the species' southern occurrence in the Cretaceous landmass Laramidia.
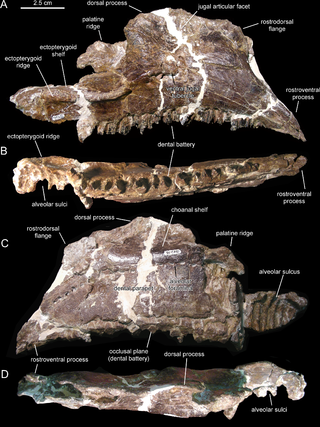
Canardia is an extinct genus of lambeosaurine dinosaur known from the Late Cretaceous Marnes d'Auzas Formation of Haute-Garonne department, in Occitanie region, southwestern France. The type species Canardia garonnensis was first described and named by Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete and Àngel Galobart in 2013. It is only known from juvenile specimens. The name of the genus comes from “canard”, the French word for “duck”, an allusion to the fact that this animal belongs to the hadrosaurids which are also known as duck-billed dinosaurs. The specific epithet garonnensis refers to the Haute-Garonne department where this dinosaur has been found. Although universally recognized as a lambeosaurine, its precise position within them is debated. Some authors consider it as a close relative of the genus Aralosaurus from Central Asia with which it would form the tribe Aralosaurini, while others include it in a more derived clade, the Arenysaurini in which all lambeosaurines from Europe and North Africa are placed. Canardia was one of the last non-avian dinosaurs and lived between 67,5 and 66 my on the former Ibero-Armorican Island, which included much of France and Spain.

This timeline of hadrosaur research is a chronological listing of events in the history of paleontology focused on the hadrosauroids, a group of herbivorous ornithopod dinosaurs popularly known as the duck-billed dinosaurs. Scientific research on hadrosaurs began in the 1850s, when Joseph Leidy described the genera Thespesius and Trachodon based on scrappy fossils discovered in the western United States. Just two years later he published a description of the much better-preserved remains of an animal from New Jersey that he named Hadrosaurus.

Adynomosaurus is a genus of lambeosaurine dinosaur from the Late Cretaceous of what is now Catalonia, Spain. First discovered in 2012, it was named in 2019 with the type and only species being Adynomosaurus arcanus. It is only known from scant material, but is distinguished from other hadrosaurs by its weakly developed shoulder blade which would have had underdeveloped musculature, which lends it its scientific name, partially from the Greek word for "weak". Its exact relationships with other hadrosaurs remain unresolved, with it not consistently being recovered as a relative of any other specific genera, though some studies have allied it with Tsintaosaurini or even found it outside of Hadrosauridae. It would have lived as part of a diverse coastal estuary ecosystem, made up of meandering rivers and mud flats. The discovery of Adynomosaurus adds to the very incomplete fossil record of hadrosaurid dinosaurs in the Late Cretaceous of Europe, and it fits into a picture of major ecological turnover that was occurring during the Maastrichtian stage in the region.

The Basturs Poble bonebed is a mega-bonebed of hadrosaur dinosaur fossils, discovered in Catalonia, Spain. Hundreds of hadrosaur fossils have been found at the site, which would have been on a large island during the Late Cretaceous when the animals preserved were alive. Despite the enormous amount of specimens, taxonomically informative material has been scarce at the site, leading to extensive debate as to its nature. The number of species present, age of the individuals present in the sample, and taxonomic identity of the remains have been the primary matters of debate. Previously considered to represent Koutalisaurus, Pararhabdodon, or multiple, perhaps dwarf species, it is currently thought that a single, indeterminate species of lambeosaurine was present at the site, and that individuals of many different ages were present.
