The Pleistocene is the geological epoch that lasted from c. 2.58 million to 11,700 years ago, spanning the Earth's most recent period of repeated glaciations. Before a change was finally confirmed in 2009 by the International Union of Geological Sciences, the cutoff of the Pleistocene and the preceding Pliocene was regarded as being 1.806 million years Before Present (BP). Publications from earlier years may use either definition of the period. The end of the Pleistocene corresponds with the end of the last glacial period and also with the end of the Paleolithic age used in archaeology. The name is a combination of Ancient Greek πλεῖστος (pleîstos), meaning "most", and καινός, meaning "new".

The Quaternary is the current and most recent of the three periods of the Cenozoic Era in the geologic time scale of the International Commission on Stratigraphy (ICS). It follows the Neogene Period and spans from 2.58 million years ago to the present. As of 2023, the Quaternary Period is divided into two epochs: the Pleistocene and the Holocene ; a third epoch, the Anthropocene, has recently been proposed, but it is not officially recognised by the ICS.

The dire wolf is an extinct canine. The dire wolf lived in the Americas during the Late Pleistocene and Early Holocene epochs. The species was named in 1858, four years after the first specimen had been found. Two subspecies are recognized: Aenocyon dirus guildayi and Aenocyon dirus dirus. The largest collection of its fossils has been obtained from the Rancho La Brea Tar Pits in Los Angeles.

The Last Glacial Period (LGP), also known colloquially as the Last Ice Age or simply Ice Age, occurred from the end of the Last Interglacial to the end of the Younger Dryas, encompassing the period c. 115,000 – c. 11,700 years ago.

The steppe bison or steppe wisent is an extinct species of bison. It was widely distributed across the mammoth steppe, ranging from Western Europe to eastern Beringia in North America during the Late Pleistocene. It is ancestral to all North American bison, including ultimately modern American bison. Three chronological subspecies, Bison priscus priscus, Bison priscus mediator, and Bison priscus gigas, have been suggested.

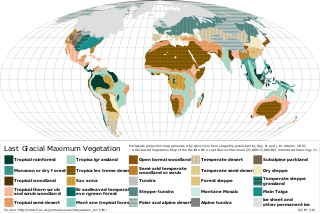
The Last Glacial Maximum (LGM), also referred to as the Last Glacial Coldest Period, was the most recent time during the Last Glacial Period where ice sheets were at their greatest extent 26,000 and 20,000 years ago. Ice sheets covered much of Northern North America, Northern Europe, and Asia and profoundly affected Earth's climate by causing a major expansion of deserts, along with a large drop in sea levels.
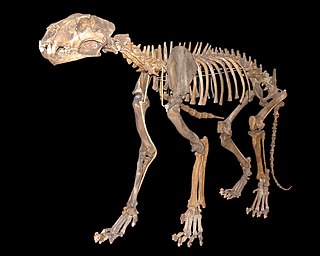
Panthera spelaea, also known as the cave lion or steppe lion, is an extinct Panthera species that most likely evolved in Europe after the third Cromerian interglacial stage, less than 600,000 years ago. Genetic analysis of ancient DNA has revealed that while closely related, it was a distinct species genetically isolated from the modern lion occurring in Africa and Asia, with the genetic divergence between the two species variously estimated between 1.9 million and 600,000 years ago. It is closely related and probably ancestral to the American lion. The species ranged from Western Europe to eastern Beringia in North America, and was a prominent member of the mammoth steppe fauna. It became extinct about 13,000 years ago.

During the Last Glacial Maximum, the mammoth steppe, also known as steppe-tundra, was once the Earth's most extensive biome. It stretched east-to-west, from the Iberian Peninsula in the west of Europe, across Eurasia to North America, through Beringia and Canada; from north-to-south, the steppe reached from the arctic islands southward to China. The mammoth steppe was cold and dry, and relatively featureless, though topography and geography varied considerably throughout. Some areas featured rivers which, through erosion, naturally created gorges, gulleys, or small glens. The continual glacial recession and advancement over millennia contributed more to the formation of larger valleys and different geographical features. Overall, however, the steppe is known to be flat and expansive grassland. The vegetation was dominated by palatable, high-productivity grasses, herbs and willow shrubs.
The Chibanian, widely known as the Middle Pleistocene, is an age in the international geologic timescale or a stage in chronostratigraphy, being a division of the Pleistocene Epoch within the ongoing Quaternary Period. The Chibanian name was officially ratified in January 2020. It is currently estimated to span the time between 0.770 Ma and 0.126 Ma, also expressed as 770–126 ka. It includes the transition in palaeoanthropology from the Lower to the Middle Paleolithic over 300 ka.

The Late Pleistocene is an unofficial age in the international geologic timescale in chronostratigraphy, also known as the Upper Pleistocene from a stratigraphic perspective. It is intended to be the fourth division of the Pleistocene Epoch within the ongoing Quaternary Period. It is currently defined as the time between c. 129,000 and c. 11,700 years ago. The late Pleistocene equates to the proposed Tarantian Age of the geologic time scale, preceded by the officially ratified Chibanian. The beginning of the Late Pleistocene is the transition between the end of the Penultimate Glacial Period and the beginning of the Last Interglacial around 130,000 years ago. The Late Pleistocene ends with the termination of the Younger Dryas, some 11,700 years ago when the Holocene Epoch began.
The Late Glacial Interstadial (LGI) c. 14,670 to c. 12,890 BP, also called the Bølling–Allerød interstadial, represents the first pronounced warming since the end of the Last Glacial Maximum (LGM). Human populations, which had previously been forced into refuge areas, gradually begin to repopulate the Northern Hemisphere's Eurasian landmass.
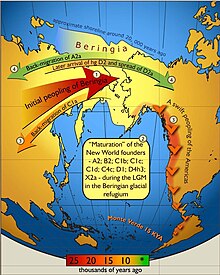
Last Glacial Maximum refugia were places (refugia) in which humans and other species survived during the Last Glacial Period, around 25,000 to 18,000 years ago. Glacial refugia are areas that climate changes were not as severe, and where species could recolonize after deglaciation.

The Late Pleistocene to the beginning of the Holocene saw numerous extinctions of predominantly megafaunal animal species, which resulted in a collapse in faunal density and diversity across the globe. The extinctions during the Late Pleistocene are differentiated from previous extinctions by the widespread absence of ecological succession to replace these extinct megafaunal species, and the regime shift of previously established faunal relationships and habitats as a consequence. The timing and severity of the extinctions varied by region and are thought to have been driven by varying combinations of human and climatic factors. Human impact on megafauna populations is thought to have been driven by hunting ("overkill"), as well as possibly environmental alteration. The relative importance of human vs climatic factors in the extinctions has been the subject of long-running controversy.

Arctodus is an extinct genus of short-faced bear that inhabited North America during the Pleistocene. There are two recognized species: the lesser short-faced bear and the giant short-faced bear. Of these species, A. simus was larger, is known from more complete remains, and is considered one of the most charismatic of North America's megafauna. A. pristinus was largely restricted to the Early Pleistocene of the Eastern United States, whereas A. simus had a broader range, with most finds being from the Late Pleistocene of the United States, Mexico and Canada. A. simus evolved from A. pristinus, but both species likely overlapped in the Middle Pleistocene. Both species are relatively rare in the fossil record.

The Beringian wolf is an extinct population of wolf that lived during the Ice Age. It inhabited what is now modern-day Alaska, Yukon, and northern British Columbia. Some of these wolves survived well into the Holocene. The Beringian wolf is an ecomorph of the gray wolf and has been comprehensively studied using a range of scientific techniques, yielding new information on their prey species and feeding behaviors. It has been determined that these wolves are morphologically distinct from modern North American wolves and genetically basal to most modern and extinct wolves. The Beringian wolf has not been assigned a subspecies classification and its relationship with the extinct European cave wolf is not clear.

The Pleistocene wolf, also referred to as the Late Pleistocene wolf, is an extinct lineage or ecomorph of the grey wolf. It was a Late Pleistocene 129 Ka – early Holocene 11 Ka hypercarnivore. While comparable in size to a large modern grey wolf, it possessed a shorter, broader palate with large carnassial teeth relative to its overall skull size, allowing it to prey and scavenge on Pleistocene megafauna. Such an adaptation is an example of phenotypic plasticity. It was once distributed across the northern Holarctic. Phylogenetic evidence indicates that despite being much smaller than this prehistoric wolf, the Japanese wolf, which went extinct in the early 20th century, was of a Pleistocene wolf lineage, thus extending its survival to several millennia after its previous estimated extinction around 7,500 years ago.
The Upward Sun River site, or Xaasaa Na’, is a Late Pleistocene archaeological site associated with the Paleo-Arctic tradition, located in the Tanana River Valley, Alaska. Dated to around 11,500 BP, Upward Sun River is the site of the oldest human remains discovered on the American side of Beringia. The site was first discovered in 2006.

The evolution of the wolf occurred over a geologic time scale of at least 300 thousand years. The grey wolf Canis lupus is a highly adaptable species that is able to exist in a range of environments and which possesses a wide distribution across the Holarctic. Studies of modern grey wolves have identified distinct sub-populations that live in close proximity to each other. This variation in sub-populations is closely linked to differences in habitat – precipitation, temperature, vegetation, and prey specialization – which affect cranio-dental plasticity.

The peopling of the Americas began when Paleolithic hunter-gatherers (Paleo-Indians) entered North America from the North Asian Mammoth steppe via the Beringia land bridge, which had formed between northeastern Siberia and western Alaska due to the lowering of sea level during the Last Glacial Maximum. These populations expanded south of the Laurentide Ice Sheet and spread rapidly southward, occupying both North and South America, by 12,000 to 14,000 years ago. The earliest populations in the Americas, before roughly 10,000 years ago, are known as Paleo-Indians. Indigenous peoples of the Americas have been linked to Siberian populations by linguistic factors, the distribution of blood types, and in genetic composition as reflected by molecular data, such as DNA.
The coastal migration hypothesis is one of two leading hypotheses about the settlement of the Americas at the time of the Last Glacial Maximum. It proposes one or more migration routes involving watercraft, via the Kurile island chain, along the coast of Beringia and the archipelagos off the Alaskan-British Columbian coast, continuing down the coast to Central and South America. The alternative is the hypothesis solely by interior routes, which assumes migration along an ice-free corridor between the Laurentide and Cordilleran ice sheets during the Last Glacial Maximum.