The ORC is present throughout the cell cycle bound to replication origins, but is only active in late mitosis and early G1.
In yeast, ORC also plays a role in the establishment of silencing at the mating-typeloci Hidden MAT Left (HML) and Hidden MAT Right (HMR).[5][6][7] ORC participates in the assembly of transcriptionally silent chromatin at HML and HMR by recruiting the Sir1 silencing protein to the HML and HMR silencers.[7][18][19]
Both Orc1 and Orc5 bind ATP, though only Orc1 has ATPase activity.[20] The binding of ATP by Orc1 is required for ORC binding to DNA and is essential for cell viability.[11] The ATPase activity of Orc1 is involved in formation of the pre-RC.[21][22][23] ATP binding by Orc5 is crucial for the stability of ORC as a whole. Only the Orc1-5 subunits are required for origin binding; Orc6 is essential for maintenance of pre-RCs once formed.[24] Interactions within ORC suggest that Orc2-3-6 may form a core complex.[4] A 2020 report suggests that budding yeast ORC dimerizes in a cell cycle dependent manner to control licensing.[25][26]
Proteins
This process was initiated by the loading of Mcm2-7 onto the chromatid by the ORC and associated proteins
The following proteins are present in the ORC:
ORC Protein Subunits, orthology and nomenclature by species[27]
Archaea feature a simplified version of the ORC, Mcm, and as a consequence the combined pre-RC. Instead of using six different mcm proteins to form a pseudo-symmetrical heterohexamer, all six subunits in the archaeal MCM are the same. They usually have multiple proteins that are homologous to both Cdc6 and Orc1, some of which perform the function of both. Unlike eukaryotic Orc, they do not always form a complex. In fact, they have divergent complex structures when these do form. Sulfolobus islandicus also uses a Cdt1 homologue to recognize one of its replication origins.[28]
Autonomously replicating sequences
Budding yeast
Autonomously Replicating Sequences (ARS), first discovered in budding yeast, are integral to the success of the ORC. These 100-200bp sequences facilitate replication activity during S phase. ARSs can be placed at any novel location of the chromosomes of budding yeast and will facilitate replication from those sites. A highly conserved sequence of 11bp (known as the A element) is thought to be essential for origin function in budding yeast.[27] The ORC was originally identified by its ability to bind to the A element of the ARS in budding yeast.
Animals
Animal cells contain a much more cryptic version of an ARS, with no conserved sequences found as of yet. Here, replication origins gather into bundles called replicon clusters. Each cluster's replicons are similar in length, but individual clusters have replicons of varying length. These replicons all have similar basic residues to which the ORC binds, which in many ways mimic the conserved 11bp A element. All of these clusters are simultaneously activated during S phase.[27]
Role in pre-RC assembly
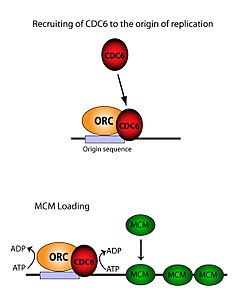
The ORC is essential for the loading of MCM complexes (Pre-RC) onto DNA. This process is dependent on the ORC, Noc3, Cdc6, and Cdt1 – involving several ATP controlled recruiting events. First, the ORC, Noc3p and Cdc6 form a complex on origin DNA (marked by ARS type regions). New ORC/Noc3/Cdc6 complexes then recruit Cdt1/Mcm2-7 molecules to the site. Once this massive ORC/Noc3/Cdc6/Cdt1/Mcm2-7 complex is formed, the ORC/Noc3/Cdc6/Cdt1 molecules work together to load Mcm2-7 onto the DNA itself by hydrolysis of ATP by Cdc6. Cdc6's phosphorylative activity is dependent on both the ORC and origin DNA. This leads to Cdt1 having decreased stability on the DNA and falling off of the complex leading to Mcm2-7 loading on to the DNA.[29][27][30][31] The structure of the ORC, MCM, as well as the intermediate OCCM complex has been resolved.[32]
Origin binding activity
Although the ORC is composed of six discrete subunits, only one of these has been found to be significant - ORC1. In vivo studies have shown that Lys-263 and Arg-367 are the basic residues responsible for faithful ORC loading. These molecules represent the above-mentioned ARS.[33]ORC1 interacts with ATP and these basic residues in order to bind the ORC to origin DNA. It has been established that this occurs far before replication, and that the ORC itself is already bound to Origin DNA by the time any Mcm2-7 loading occurs.[31] When Mcm2-7 is first loaded it completely encircles the DNA and helicase activity is inhibited. In S phase, the Mcm2-7 complex interacts with helicase cofactorsCdc45 and GINS to isolate a single DNA strand, unwind the origin, and begin replication down the chromosome. In order to have bidirectional replication, this process happens twice at an origin. Both loading events are mediated by one ORC via an identical process as the first.[34]
↑ Dutta A, Bell SP (1997). "Initiation of DNA replication in eukaryotic cells". Annual Review of Cell and Developmental Biology. 13: 293–332. doi:10.1146/annurev.cellbio.13.1.293. PMID9442876.
1 2 3 Matsuda K, Makise M, Sueyasu Y, Takehara M, Asano T, Mizushima T (December 2007). "Yeast two-hybrid analysis of the origin recognition complex of Saccharomyces cerevisiae: interaction between subunits and identification of binding proteins". FEMS Yeast Research. 7 (8): 1263–9. doi:10.1111/j.1567-1364.2007.00298.x. PMID17825065.
↑ Fox CA, Ehrenhofer-Murray AE, Loo S, Rine J (June 1997). "The origin recognition complex, SIR1, and the S phase requirement for silencing". Science. 276 (5318): 1547–51. doi:10.1126/science.276.5318.1547. PMID9171055.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.