
Gracilisuchus is an extinct genus of tiny pseudosuchian from the Late Triassic of Argentina. It contains a single species, G. stipanicicorum, which is placed in the clade Suchia, close to the ancestry of crocodylomorphs. Both the genus and the species were first described by Alfred Romer in 1972.

Doswellia is an extinct genus of archosauriform from the Late Triassic of North America. It is the most notable member of the family Doswelliidae, related to the proterochampsids. Doswellia was a low and heavily built carnivore which lived during the Carnian stage of the Late Triassic. It possesses many unusual features including a wide, flattened head with narrow jaws and a box-like rib cage surrounded by many rows of bony plates. The type species Doswellia kaltenbachi was named in 1980 from fossils found within the Vinita member of the Doswell Formation in Virginia. The formation, which is found in the Taylorsville Basin, is part of the larger Newark Supergroup. Doswellia is named after Doswell, the town from which much of the taxon's remains have been found. A second species, D. sixmilensis, was described in 2012 from the Bluewater Creek Formation of the Chinle Group in New Mexico; however, this species was subsequently transferred to a separate doswelliid genus, Rugarhynchos. Bonafide Doswellia kaltenbachi fossils are also known from the Chinle Formation of Arizona.

Proterochampsidae is a family of proterochampsian archosauriforms. Proterochampsids may have filled an ecological niche similar to modern crocodiles, and had a general crocodile-like appearance. They lived in what is now South America in the Middle and Late Triassic.
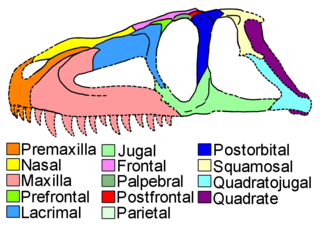
Luperosuchus is an extinct genus of loricatan pseudosuchian reptile which contains only a single species, Luperosuchus fractus. It is known from the Chañares Formation of Argentina, within strata belonging to the latest Ladinian stage of the late Middle Triassic, or the earliest Carnian of the Late Triassic. Luperosuchus was one of the largest carnivores of the Chañares Formation, although its remains are fragmentary and primarily represented by a skull with similarities to Prestosuchus and Saurosuchus.
Yarasuchus is an extinct genus of avemetatarsalian archosaur that lived during the Anisian stage of the Middle Triassic of India. The genus was named and described in 2005 from a collection of disarticulated but fairly complete fossil material found from the Middle Triassic Yerrapalli Formation. The material is thought to be from two individuals, possibly three, with one being much more complete and articulated than the other. The type and only species is Y. deccanensis. Yarasuchus was a quadruped roughly 2–2.5 metres (6.6–8.2 ft) long, with an elongated neck and tall spines on its vertebrae. Unlike other quadrupedal Triassic reptiles, the limbs and shoulders of Yarasuchus were slender, and more like those of ornithodirans.

Vancleavea is a genus of extinct, armoured, non-archosaurian archosauriforms from the Late Triassic of western North America. The type and only known species is V. campi, named by Robert Long & Phillip A Murry in 1995. At that time, the genus was only known from fragmentary bones including osteoderms and vertebrae. However, since then many more fossils have been found, including a pair of nearly complete skeletons discovered in 2002. These finds have shown that members of the genus were bizarre semiaquatic reptiles. Vancleavea individuals had short snouts with large, fang-like teeth, and long bodies with small limbs. They were completely covered with bony plates known as osteoderms, which came in several different varieties distributed around the body. Phylogenetic analyses by professional paleontologists have shown that Vancleavea was an archosauriform, part of the lineage of reptiles that would lead to archosaurs such as dinosaurs and crocodilians. Vancleavea lacks certain traits which are present in most other archosauriforms, most notably the antorbital, mandibular and supratemporal fenestrae, which are weight-saving holes in the skulls of other taxa. However, other features clearly support its archosauriform identity, including a lack of intercentra, the presence of osteoderms, an ossified laterosphenoid, and several adaptations of the femur and ankle bones. In 2016, a new genus of archosauriform, Litorosuchus, was described. This genus resembled both Vancleavea and more typical archosauriforms in different respects, allowing Litorosuchus to act as a transitional fossil linking Vancleavea to less aberrant archosauriforms.
Koilamasuchus is an extinct genus of indeterminate archosauriform from the Triassic of Argentina. It is based on an external mold of a partial postcranial skeleton from the Quebrada de los Fósiles Formation. Due to its incomplete nature, the relationships of this reptile are difficult to establish. Originally described as a non-archosaur archosauriform, later studies tentatively considered it a doswelliid or a suchian archosaur.

Tarjadia is an extinct genus of erpetosuchid pseudosuchian, distantly related to modern crocodilians. It is known from a single species, T. ruthae, first described in 1998 from the Middle Triassic Chañares Formation in Argentina. Partial remains have been found from deposits that are Anisian-Ladinian in age. Long known mostly from osteoderms, vertebrae, and fragments of the skull, specimens described in 2017 provided much more anatomical details and showed that it was a fairly large predator. Tarjadia predates known species of aetosaurs and phytosaurs, two Late Triassic groups of crurotarsans with heavy plating, making it one of the first heavily armored archosaurs. Prior to 2017, most studies placed it outside Archosauria as a member of Doswelliidae, a family of heavily armored and crocodile-like archosauriforms. The 2017 specimens instead show that it belonged to the Erpetosuchidae.

Rhadinosuchus is an extinct genus of proterochampsian archosauriform reptile from the Late Triassic. It is known only from the type species Rhadinosuchus gracilis, reposited in Munich, Germany. The fossil includes an incomplete skull and fragments of post-cranial material. Hosffstetter (1955), Kuhn (1966), Reig (1970) and Bonaparte (1971) hypothesized it to be synonymous with Cerritosaurus, but other characteristics suggest it is closer to Chanaresuchus and Gualosuchus, while it is certainly different from Proterochampsa and Barberenachampsa. The small size indicates it is a young animal, making it hard to classify.
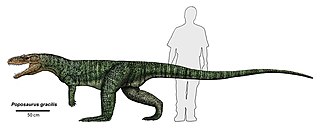
Poposauroidea is a clade of advanced pseudosuchians. It includes poposaurids, shuvosaurids, ctenosauriscids, and other unusual pseudosuchians such as Qianosuchus and Lotosaurus. It excludes most large predatory quadrupedal "rauisuchians" such as rauisuchids and "prestosuchids". Those reptiles are now allied with crocodylomorphs in a clade known as Loricata, which is the sister taxon to the poposauroids in the clade Paracrocodylomorpha. Although it was first formally defined in 2007, the name "Poposauroidea" has been used for many years. The group has been referred to as Poposauridae by some authors, although this name is often used more narrowly to refer to the family that includes Poposaurus and its close relatives.
Doswelliidae is an extinct family of carnivorous archosauriform reptiles that lived in North America and Europe during the Middle to Late Triassic period. Long represented solely by the heavily-armored reptile Doswellia, the family's composition has expanded since 2011, although two supposed South American doswelliids were later redescribed as erpetosuchids. Doswelliids were not true archosaurs, but they were close relatives and some studies have considered them among the most derived non-archosaurian archosauriforms. They may have also been related to the Proterochampsidae, a South American family of crocodile-like archosauriforms.
Proterochampsia is a clade of early archosauriform reptiles from the Triassic period. It includes the Proterochampsidae and probably also the Doswelliidae. Nesbitt (2011) defines Proterochampsia as a stem-based taxon that includes Proterochampsa and all forms more closely related to it than Euparkeria, Erythrosuchus, Passer domesticus, or Crocodylus niloticus. Therefore, the inclusion of Doswelliidae in it is dependent upon whether Doswellia and Proterochampsa form a monophyletic group to the exclusion of Archosauria and other related groups.
Aetobarbakinoides is an extinct genus of stagonolepidid aetosaur known from the Late Triassic of Rio Grande do Sul state, southern Brazil. Fossils have been found from the Santa Maria Supersequence of the late Carnian and early Norian stages, making Aetobarbakinoides one of the oldest aetosaurs. The type species, A. brasiliensis, was named in 2012, and is notable for being described primarily by features of the vertebrae; most other aetosaurs are diagnosed by features in bony plates called osteoderms, which are by far the most common material. Although placed in a basal phylogenetic position among aetosaurs, Aetobarbakinoides is closely related to both desmatosuchines and typothoracisines, two derived clades of aetosaurs.

Erpetosuchidae is an extinct family of pseudosuchian archosaurs. Erpetosuchidae was named by D. M. S. Watson in 1917 to include Erpetosuchus. It includes the type species Erpetosuchus granti from the Late Triassic of Scotland, Erpetosuchus sp. from the Late Triassic of eastern United States and Parringtonia gracilis from the middle Middle Triassic of Tanzania; the group might also include Dyoplax arenaceus from the Late Triassic of Germany, Archeopelta arborensis and Pagosvenator candelariensis from Brazil and Tarjadia ruthae from Argentina.
Jaxtasuchus is an extinct genus of armored doswelliid archosauriform reptile known from the Middle Triassic of the Erfurt Formation in Germany. The type species, Jaxtasuchus salomoni, was named in 2013 on the basis of several incomplete skeletons and other isolated remains. Like other doswelliids, members of the genus were heavily armored, with four longitudinal rows of bony plates called osteoderms covering the body. Jaxtasuchus is the first doswelliid known from Europe and is most closely related to Doswellia from the Late Triassic of the eastern United States. However, it was not as specialized as Doswellia, retaining several generalized archosauriform characteristics and having less armor. Jaxtasuchus fossils have been found in aquatic mudstones alongside fossils of temnospondyl amphibians, crustaceans, and mollusks, suggesting that Jaxtasuchus was semiaquatic like modern crocodilians.
Nundasuchus is an extinct genus of crurotarsan, possibly a suchian archosaur related to Paracrocodylomorpha. Remains of this genus are known from the Middle Triassic Manda beds of southwestern Tanzania. It contains a single species, Nundasuchus songeaensis, known from a single partially complete skeleton, including vertebrae, limb elements, osteoderms, and skull fragments.
Aenigmaspina is an extinct genus of enigmatic pseudosuchian (=crurotarsan) archosaur from the Late Triassic of the United Kingdom. Its fossils are known from the Pant-y-ffynnon Quarry in South Wales, of which its type and only known species is named after, A. pantyffynnonensis. Aenigmaspina is characterised by the unusual spines on its vertebrae, which are broad and flat on top with a unique 'Y' shape. Although parts of its skeleton is relatively well known, the affinities of Aenigmaspina to other pseudosuchians are unclear, although it is possibly related to families Ornithosuchidae, Erpetosuchidae or Gracilisuchidae.

Dynamosuchus is an extinct genus of pseudosuchian archosaurs from the family Ornithosuchidae. It is known from a single species, Dynamosuchus collisensis, which is based on a partial skeleton from the Santa Maria Formation of Brazil. Dynamosuchus is considered a close relative of Venaticosuchus, which is known from the Ischigualasto Formation of Argentina. Ornithosuchids are one of many groups which lived in the Santa Maria and Ischigualasto Formations, which formed at approximately the same time and were ecologically similar. As a large scavenging reptile, Dynamosuchus helps to illuminate the trophic structure of the Santa Maria Formation. It also supports the hypothesis that ornithosuchids had diversified throughout South America by the start of the Carnian, and were not originally endemic to the Ischigualasto-Villa Unión Basin.

Rugarhynchos is an extinct genus of doswelliid archosauriform from the Late Triassic of New Mexico. The only known species is Rugarhynchos sixmilensis. It was originally described as a species of Doswellia in 2012, before receiving its own genus in 2020. Rugarhynchos was a close relative of Doswellia and shared several features with it, such as the absence of an infratemporal fenestra and heavily textured skull bones. However, it could also be distinguished by many unique characteristics, such as a thick diagonal ridge on the side of the snout, blunt spikes on its osteoderms, and a complex suture between the quadratojugal, squamosal, and jugal. Non-metric multidimensional scaling and tooth morphology suggest that Rugarhynchos had a general skull anatomy convergent with some crocodyliforms, spinosaurids, and phytosaurs. However, its snout was somewhat less elongated than those other reptiles.

Polymorphodon is an extinct genus of archosauriform reptile from the Middle Triassic of Germany. The only known species is Polymorphodon adorfi, discovered in Lower Keuper deposits at a quarry in Eschenau, Germany. Polymorphodon is notable for its heterodont dentition, with long and conical premaxillary teeth followed by thin maxillary teeth with large serrations. Maxillary teeth near the back of the mouth are short and leaf-shaped, similar to some living and extinct reptiles with a herbivorous or omnivorous diet. This may suggest that Polymorphodon had some reliance on plants in its diet, a rarity among basal archosauriforms, most of which are carnivores.



