| Genus | Authority | Year | Location | Unit | Age | Traditional classification | Modern classification | Notes | Images |
|---|
| Apatosuchus | Sues & Schoch | 2013 |  Germany Germany | Löwenstein Formation (Stubensandstein) | Norian | Theropoda | Loricata | A small "rauisuchian" based on a partial skull. Originally considered a species of Halticosaurus , a contemporary theropod dinosaur. | |
| Arganasuchus | Jalil & Peyer | 2007 |  Morocco Morocco | Timezgadiouine Formation | Carnian? | Rauisuchidae | Loricata | A "rauisuchian" based on scant skull and postcranial material similar to that of Batrachotomus , Luperosuchus , and Postosuchus . |  |
| Arizonasaurus | Welles | 1947 |  United States United States | Moenkopi Formation | Anisian | Theropoda? Rauisuchia? | Poposauroidea (Ctenosauriscidae) | One of the most complete ctenosauriscids, owing to a partial skeleton with skull and hip material discovered in 2002. Vital for understanding the affinities of ctenosauriscids with poposaurs. |  |
| Batrachotomus | Gower | 1999 | Germany | Erfurt Formation (Lower Keuper) | Late Ladinian | Prestosuchidae / Rauisuchidae | Loricata | One of the most completely-known "rauisuchians", with numerous fossils recovered from sites at Kupferzell, Crailsheim and Vellberg-Eschenau. | 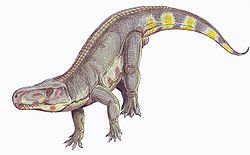 |
| Bromsgroveia | Galton | 1985 |  United Kingdom United Kingdom | Bromsgrove Sandstone | Anisian | Theropoda / Temnospondyli /Poposauridae | Poposauroidea (Ctenosauriscidae) | Named based on a distinctive poposaur-like ilium, but also possibly incorporating "rauisuchian" fossil material such as teeth and vertebrae described from England since the mid-19th century. | |
| Bystrowisuchus | Sennikov | 2012 |  Russia Russia | Lipovskaya Formation (Gamskian Gorizont) | Late Olenekian? | Ctenosauriscidae | Poposauroidea (Ctenosauriscidae) | An early ctenosauriscid with low-spined cervical (neck) vertebrae similar to Xilousuchus . | |
| Ctenosauriscus | Kuhn | 1964 | Germany | Solling Formation | Late Olenekian to Anisian? | "Pelycosauria" / Temnospondyli / Ctenosauriscidae | Poposauroidea (Ctenosauriscidae) | Known from slabs of sail-backed dorsal vertebrae first discovered in 1871 and originally named as the (preoccupied) genus Ctenosaurus in 1902. Its affinities were strongly debated until stabilizing as poposauroid archosaur upon the discovery of new Arizonasaurus and Lotosaurus fossils. |  |
| Dagasuchus | Lacerda et al. | 2015 |  Brazil Brazil | Middle Santa Maria Formation ( Santacruzodon Assemblage Zone) | Late Ladinian or early Carnian | N/A | Loricata | A medium-sized "rauisuchian" based on a hip bone which helps to fill a gap in the biostratigraphic record of Brazilian archosaurs. | |
| Decuriasuchus | França | 2011 | Brazil | Lower Santa Maria Formation ( Dinodontosaurus Assemblage Zone) | Late Ladinian or early Carnian | Prestosuchidae | Loricata | A medium-sized "rauisuchian" named from 10 individuals (including several nearly-complete skeletons) which died and fossilized together, suggesting a gregarious social structure. |  |
| Effigia | Nesbitt & Norell | 2006 | United States | Chinle Formation ("siltstone member") | Late Norian or Rhaetian | Derived Suchia | Poposauroidea (Shuvosauridae) | A bizarre pseudosuchian convergent on ornithomimids, with a toothless skull, theropod-like hip, and very short arms. Discovered within a sediment block collected from the Coelophysis Quarry of Ghost Ranch, New Mexico. |  |
| Etjosuchus | Tolchard et al. | 2021 |  Namibia Namibia | Omingonde Formation | Ladinian or early Carnian | Erythrosuchidae | Loricata | A large and possibly bipedal "rauisuchian" based on a partial skeleton previously misattributed to the erythrosuchid Erythrosuchus . | |
| Fasolasuchus | Bonaparte | 1981 |  Argentina Argentina | Los Colorados Formation | Late Norian | Rauisuchidae | Loricata | One of the last and largest "rauisuchians", occurring alongside an increasingly diverse fauna of sauropodomorph dinosaurs. |  |
| Heptasuchus | Dawley et al. | 1979 | United States | Popo Agie Formation | Late Carnian? | Prestosuchidae? | Loricata | One of the oldest predatory archosaurs from North America, based on skull and postcranial fragments from Wyoming. Probably a close relative of Batrachotomus . |  |
| Hypselorhachis | Butler et al. | 2009 |  Tanzania Tanzania | Manda Formation | Anisian? to Carnian? | Ctenosauriscidae | Poposauroidea (Ctenosauriscidae) | A ctenosauriscid based on a single well-preserved vertebra with a very long neural spine. First mentioned in an unpublished 1966 by Alan Charig, but not formally described until 2009. | 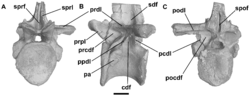 |
| Lotosaurus | Zhang | 1975 |  China China | Badong Formation | Anisian? to Carnian? | "Lotosauridae" | Poposauroidea | An unusual quadruped combining a neural spine sail with a toothless skull. Known from abundant fossils clustered into a bonebed, but most of these fossils remain undescribed. | 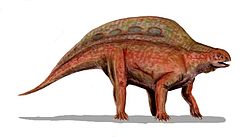 |
| Luperosuchus | Romer | 1971 | Argentina | Chanares Formation ( Tarjadia Assemblage Zone) | Late Ladinian or earliest Carnian | Prestosuchidae | Loricata (Prestosuchidae?) | A medium-sized "rauisuchian" with a skull similar to Prestosuchus and Saurosuchus . |  |
| Mambawakale | Butler et al. | 2022 | Tanzania | Manda Formation | Anisian? to Carnian? | "Pallisteriidae" | Basal Paracrocodylomorpha? | A possible paracrocodylomorph based on a massive partial skull. Mentioned as "Pallisteria angustimentum" in an unpublished 1967 manuscript by Alan Charig, but not formally described (and provided a new scientific name) until 2022. |  |
| Mandasuchus | Butler et al. | 2018 | Tanzania | Manda Formation | Anisian? to Carnian? | Prestosuchidae? | Basal Paracrocodylomorpha? | One of the most basal loricatans or poposauroids, first mentioned in a 1956 doctoral thesis by Alan Charig but not formally described until 2018. | |
| Polonosuchus | Brusatte et al. | 2009 |  Poland Poland | Krasiejów claypit | Late Carnian? | Rauisuchidae | Loricata (Rauisuchidae) | A large rauisuchid, formerly named as a species of Teratosaurus and currently considered a close relative of Postosuchus . |  |
| Poposaurus | Mehl | 1915 | United States | Popo Agie Formation, Dockum Group, Chinle Formation (Monitor Butte Member, Blue Mesa Member, Sonsela Member) | Late Carnian? to middle Norian | Dinosauria / Poposauridae | Poposauroidea (Poposauridae) | A widespread bipedal carnivore with hip bones so similar to dinosaurs that for decades it was mistaken as one. Though skull fragments are very rare, the postcrania is well-described according to a nearly complete skeleton discovered in Utah in 2003. |  |
| Postosuchus | Chatterjee | 1985 | United States | Cooper Canyon Formation, Chatham Group?, Chinle Formation? | Norian | Poposauridae / Rauisuchidae | Loricata (Rauisuchidae) | A large rauisuchid with a short skull and possibly bipedal locomotion. One of the most well-described and widespread "rauisuchians" from North America, though not all fossils referred to the genus may actually belong to it. Proposed as an ancestor to tyrannosaurids when first described. |  |
| Prestosuchus | Huene | 1942 | Brazil | Lower Santa Maria Formation ( Dinodontosaurus Assemblage Zone) | Late Ladinian or early Carnian | Prestosuchidae | Loricata (Prestosuchidae) | Among the largest and most well-described "rauisuchians", a quadrupedal form based on multiple partial skeletons. One massive well-preserved skull has been named under the dubious genus "Karamuru vorax". |  |
| Procerosuchus | Huene | 1938 | Brazil | Lower Santa Maria Formation ( Dinodontosaurus Assemblage Zone) | Late Ladinian or early Carnian | Prestosuchidae | Loricata (Prestosuchidae?) | A poorly-known taxon, possibly a synonym or species of Prestosuchus. | |
| Qianosuchus | Li et al. | 2006 | China | Guanling Formation (Panxian biota) | Anisian | Indeterminate Crurotarsi | Poposauroidea | A small, gracile archosaur, the only known "rauisuchians" with semiaquatic and piscivorous habits owing to its narrow skull and tall tail vertebrae. |  |
| Rauisuchus | Huene | 1942 | Brazil | Upper Santa Maria Formation ( Hyperodapedon Assemblage Zone) | Middle-late Carnian | Rauisuchidae | Loricata (Rauisuchidae) | The namesake of Rauisuchia. Known primarily from skull, vertebral, and hindlimb fossils, which reconstruct a stocky quadrupedal predator with a boxy skull full of serrated teeth. |  |
| Saurosuchus | Reig | 1959 | Argentina | Ischigualasto Formation | Late Carnian | Prestosuchidae | Loricata (Prestosuchidae?) | A very large quadruped which lived alongside some of the earliest dinosaurs. Fossils include a well-preserved cranium. |  |
| Scolotosuchus | Sennikov | 2023 | Russia | Lipovskaya Formation | Late Olenekian | Rauisuchidae | "Rauisuchidae" | A large rauisuchian based on distinctive vertebrae. | |
| Shuvosaurus | Chatterjee | 1993 | United States | Cooper Canyon Formation | Early-middle Norian | Theropoda / "Chatterjeeidae" | Poposauroidea (Shuvosauridae) | An unusual beaked biped with two sets of fossils independently discovered in the mid-1990s: a toothless skull (Shuvosaurus) attributed to an ornithomimosaur-like dinosaur, and postcranial material ("Chatterjeea") comparable to poposaurids. The synonymy between these two fossil sets would not be confirmed until the discovery of Effigia, a close relative. |  |
| Sillosuchus | Alcober & Parrish | 1997 | Argentina | Ischigualasto Formation | Late Carnian | Poposauridae | Poposauroidea (Shuvosauridae) | A probable relative of Effigia and Shuvosaurus based on its vertebrae and hip fossils. May have reached up to 10 meters (33 ft) in length according to a large isolated vertebra. |  |
| Stagonosuchus | Huene | 1938 | Tanzania | Manda Formation | Anisian? to Carnian? | Prestosuchidae | Loricata | A Tanzanian archosaur named from hip and vertebral fragments. May be a species of Prestosuchus. | |
| Teratosaurus | Meyer | 1861 | Germany | Löwenstein Formation (Stubensandstein) | Norian | Carnosauria / Rauisuchidae / Poposauridae | Loricata (Rauisuchidae?) | The earliest "rauisuchian" to be named, though its holotype fossil (a toothed maxilla bone) was misidentified as that of an early carnosaurian dinosaur. A complete re-evaluation of archosaur systematics in the 1980s involved the recognition that Teratosaurus (and other "rauisuchians") were completely unrelated to carnosaurian dinosaurs. |  |
| Ticinosuchus | Krebs | 1965 |  Switzerland, Switzerland,  Italy Italy
| Mittlere Grenzbitumenzone | Late Anisian | Prestosuchidae | Suchia (non-paracrocodylomorph) | Described from a flattened but complete skeleton from Monte San Giorgio on the Swiss-Italian border. Its discovery established knowledge of "rauisuchian" proportions and their association with chirothere footprints. Commonly considered the sister taxon to Paracrocodylomorpha. |  |
| Tikisuchus | Chatterjee & Majumdar | 1987 |  India India | Tiki Formation | Carnian? | Rauisuchidae | Loricata (Rauisuchidae?) | A medium-sized probable rauisuchid with a proportionally large skull. | |
| Vivaron | Lessner et al. | 2016 | United States | Chinle Formation (Petrified Forest Member) | Middle Norian | N/A | Loricata (Rauisuchidae) | Known from Postosuchus -like skull and hip fragments from the Hayden Quarry at Ghost Ranch, New Mexico. Its discovery casts doubt on the assumption that all rauisuchid fossils from the southwestern United States can be referred to Postosuchus. |  |
| Xilousuchus | Wu | 1981 | China | Heshanggou Formation | Late Olenekian or early Anisian | Proterosuchidae / Erythrosuchidae | Poposauroidea (Ctenosauriscidae) | One of the better-understood early "rauisuchians", a ctenosauriscid including skull, braincase, and cervical (neck) fossils. | |
| Youngosuchus | Young | 1973 | China | Ermaying Formation | Anisian? | Erythrosuchidae /Rauisuchidae? | Loricata | Based on a well-preserved but poorly-described partial skeleton, including a skull. Originally named as a species of Vjushkovia , an erythrosuchid which has subsequently been synonymized with Garjainia . |  |
|




























