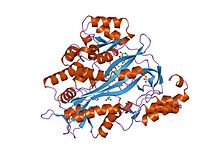
Glutathione synthetase (GSS) (EC6.3.2.3) is the second enzyme in the glutathione (GSH) biosynthesis pathway. It catalyses the condensation of gamma-glutamylcysteine and glycine, to form glutathione.[2] Glutathione synthetase is also a potent antioxidant. It is found in many species including bacteria, yeast, mammals, and plants.[3]
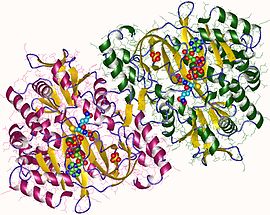
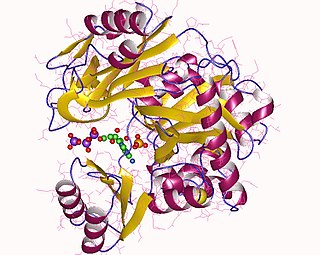
Human and yeast glutathione synthetases are homodimers, meaning they are composed of two identical subunits of itself non-covalently bound to each other. On the other hand, E. coli glutathione synthetase is a homotetramer.[3] Nevertheless, they are part of the ATP-graspsuperfamily, which consists of 21 enzymes that contain an ATP-grasp fold.[7] Each subunit interacts with each other through alpha helix and beta sheethydrogen bonding interactions and contains two domains. One domain facilitates the ATP-grasp mechanism[8] and the other is the catalytic active site for γ-glutamylcysteine. The ATP-grasp fold is conserved within the ATP-grasp superfamily and is characterized by two alpha helices and beta sheets that hold onto the ATP molecule between them.[9] The domain containing the active site exhibits interesting properties of specificity. In contrast to γ-glutamylcysteine synthetase, glutathione synthetase accepts a large variety of glutamyl-modified analogs of γ-glutamylcysteine, but is much more specific for cysteine-modified analogs of γ-glutamylcysteine.[10] Crystalline structures have shown glutathione synthetase bound to GSH, ADP, two magnesium ions, and a sulfate ion.[11] Two magnesium ions function to stabilize the acylphosphate intermediate, facilitate binding of ATP, and activate removal of phosphate group from ATP. Sulfate ion serves as a replacement for inorganic phosphate once the acylphosphate intermediate is formed inside the active site.[6]
Key residues that interact with ATP near the active site. Magnesium ions are shown in black. Generated from 2HGS.
This enzyme belongs to the family of ligases, specifically those forming carbon-nitrogen bonds as acid-D-amino-acid ligases (peptide synthases). The systematic name of this enzyme class is gamma-L-glutamyl-L-cysteine:glycine ligase (ADP-forming). Other names in common use include glutathione synthetase, and GSH synthetase. This enzyme participates in glutamate metabolism and glutathione metabolism. At least one compound, Phosphinate is known to inhibit this enzyme.[citation needed]
The biosynthetic mechanisms for synthetases use energy from nucleoside triphosphates, whereas synthases do not.[12] Glutathione synthetase stays true to this rule, in that it uses the energy generated by ATP. Initially, the carboxylate group on γ-glutamylcysteine is converted into an acyl phosphate by the transfer of an inorganic phosphate group of ATP to generate an acyl phosphate intermediate. Then the amino group of glycine participates in a nucleophilic attack, displacing the phosphate group and forming GSH.[13] After the final GSH product is made, it can be used by glutathione peroxidase to neutralize reactive oxygen species (ROS) such as H2O2 or Glutathione S-transferases in the detoxification of xenobiotics.[7]
Reaction mechanism for GSH biosynthesis. Glutamate and cysteine side chains are shown in red and green, respectively.
Function
Glutathione synthetase is important for a variety of biological functions in multiple organisms. In Arabidopsis thaliana, low levels of glutathione synthetase have resulted in increased vulnerability to stressors such as heavy metals, toxic organic chemicals, and oxidative stress.[15] The presence of a thiol functional group allows its product GSH to serve both as an effective oxidizing and reducing agent in numerous biological scenarios. Thiols can easily accept a pair of electrons and become oxidized to disulfides, and the disulfides can be readily reduced to regenerate thiols. Additionally, the thiol side chain of cysteines serve as potent nucleophiles and react with oxidants and electrophilic species that would otherwise cause damage to the cell.[16] Interactions with certain metals also stabilize thiolate intermediates.[17]
In humans, glutathione synthetase functions in a similar manner. Its product GSH participates in cellular pathways involved in homeostasis and cellular maintenance. For instance, glutathione peroxidases catalyze the oxidation of GSH to glutathione disulfide (GSSG) by reducing free radicals and reactive oxygen species such as hydrogen peroxide.[18]Glutathione S-transferase uses GSH to clean up various metabolites, xenobiotics, and electrophiles to mercapturates for excretion.[19] Because of its antioxidant role, GSS mostly produce GSH inside the cytoplasm of liver cells and imported to mitochondria where detoxification occurs.[20] GSH is also essential for the activation of the immune system to generate robust defense mechanisms against invading pathogens.[19] GSH is capable of preventing infection from the influenza virus.[21][22]
Glutathione (GSH) is synthesized in the cytoplasm of liver cells and imported to the mitochondria where it functions as a cofactor for a number of antioxidant and detoxifying enzymes.
Clinical significance
Patients with mutations in the GSS gene develop glutathione synthetase (GSS) deficiency, an autosomal recessive disorder.[23] Patients develop a wide range of symptoms depending on the severity of the mutations. Mildly affected patients experience a compensated haemolytic anaemia because mutations affect stability of the enzyme. Moderately and severely affected individuals have enzymes with dysfunctional catalytic sites, rendering it unable to participate in detoxification reactions. Physiological symptoms include metabolic acidosis, neurological defects, and increased susceptibility to pathogenic infections.[4]
Treatment of individuals with glutathione synthetase deficiency generally involve therapeutic treatments to address mild to severe symptoms and conditions. In order to treat metabolic acidosis, severely affected patients are given large amounts of bicarbonate and antioxidants such as vitamin E and vitamin C.[24] In mild cases, ascorbate and N-acetylcysteine have been shown to increase glutathione levels and increase erythrocyte production.[25] It is important to note that because glutathione synthetase deficiency is so rare, it is poorly understood. The disease also appears on a spectrum, so it is even more difficult to generalize among the few cases that occur.[26]
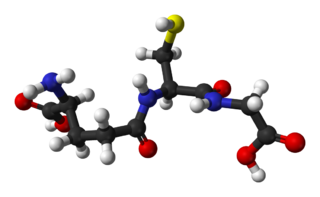
Glutathione is an organic compound with the chemical formula HOCOCH(NH2)CH2CH2CONHCH(CH2SH)CONHCH2COOH. It is an antioxidant in plants, animals, fungi, and some bacteria and archaea. Glutathione is capable of preventing damage to important cellular components caused by sources such as reactive oxygen species, free radicals, peroxides, lipid peroxides, and heavy metals. It is a tripeptide with a gamma peptide linkage between the carboxyl group of the glutamate side chain and cysteine. The carboxyl group of the cysteine residue is attached by normal peptide linkage to glycine.
Succinyl coenzyme A synthetase is an enzyme that catalyzes the reversible reaction of succinyl-CoA to succinate. The enzyme facilitates the coupling of this reaction to the formation of a nucleoside triphosphate molecule from an inorganic phosphate molecule and a nucleoside diphosphate molecule. It plays a key role as one of the catalysts involved in the citric acid cycle, a central pathway in cellular metabolism, and it is located within the mitochondrial matrix of a cell.
Sulfur assimilation is the process by which living organisms incorporate sulfur into their biological molecules. In plants, sulfate is absorbed by the roots and then transported to the chloroplasts by the transipration stream where the sulfur are reduced to sulfide with the help of a series of enzymatic reactions. Furthermore, the reduced sulfur is incorporated into cysteine, an amino acid that is a precursor to many other sulfur-containing compounds. In animals, sulfur assimilation occurs primarily through the diet, as animals cannot produce sulfur-containing compounds directly. Sulfur is incorporated into amino acids such as cysteine and methionine, which are used to build proteins and other important molecules.
Glutathione synthetase deficiency (GSD) is a rare autosomal recessive metabolic disorder that prevents the production of glutathione. Glutathione helps prevent damage to cells by neutralizing harmful molecules generated during energy production. Glutathione also plays a role in processing medications and cancer-causing compounds (carcinogens), and building DNA, proteins, and other important cellular components.
Carnosine synthase is an enzyme that catalyzes the chemical reaction
In enzymology, a D-alanine—D-alanine ligase is an enzyme that catalyzes the chemical reaction
In enzymology, a dihydrofolate synthase is an enzyme that catalyzes the chemical reaction
Glutamate–cysteine ligase (GCL) EC 6.3.2.2), previously known as γ-glutamylcysteine synthetase (GCS), is the first enzyme of the cellular glutathione (GSH) biosynthetic pathway that catalyzes the chemical reaction:
Glu-tRNAGln amidotransferase or glutaminyl-tRNA synthase (glutamine-hydrolysing) enzyme is an amidotransferase that catalyzes the conversion of the non-cognate amino acid glutamyl-tRNAGln to the cognate glutaminyl-tRNAGln.. It catalyzes the reaction:
In enzymology, a glutathionylspermidine synthase is an enzyme that catalyzes the chemical reaction
In enzymology, a homoglutathione synthase is an enzyme that catalyzes the chemical reaction
Phosphoribosylamine—glycine ligase, also known as glycinamide ribonucleotide synthetase (GARS), (EC 6.3.4.13) is an enzyme that catalyzes the chemical reaction
In enzymology, a trypanothione synthase (EC 6.3.1.9) is an enzyme that catalyzes the chemical reaction
In enzymology, a UDP-N-acetylmuramoyl-tripeptide—D-alanyl-D-alanine ligase is an enzyme that catalyzes the chemical reaction
Glutamate–cysteine ligase catalytic subunit is an enzyme that in humans is encoded by the GCLC gene.
Glutamate-cysteine ligase regulatory subunit is an enzyme that in humans is encoded by the GCLM gene.
γ-L-Glutamyl-L-cysteine, also known as γ-glutamylcysteine (GGC), is a dipeptide found in animals, plants, fungi, some bacteria, and archaea. It has a relatively unusual γ-bond between the constituent amino acids, L-glutamic acid and L-cysteine and is a key intermediate in the γ-glutamyl cycle first described by Meister in the 1970s. It is the most immediate precursor to the antioxidant glutathione.
Coenzyme gamma-F420-2:α-L-glutamate ligase (EC 6.3.2.32, MJ1001, CofF protein, gamma-F420-2:alpha-L-glutamate ligase) is an enzyme with systematic name L-glutamate:coenzyme gamma-F420-2 (ADP-forming). This enzyme catalyses the following chemical reaction
In molecular biology, the ATP-grasp fold is a unique ATP-binding protein structural motif made of two α+β subdomains that "grasp" a molecule of ATP between them. ATP-grasp proteins have ATP-dependent carboxylate-amine/thiol ligase activity.
↑ Hara T, Kato H, Katsube Y, Oda J (Sep 1996). "A pseudo-michaelis quaternary complex in the reverse reaction of a ligase: structure of Escherichia coli B glutathione synthetase complexed with ADP, glutathione, and sulfate at 2.0 A resolution". Biochemistry. 35 (37): 11967–74. doi:10.1021/bi9605245. PMID8810901.
↑ "Synthases and Ligases". IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN), and Nomenclature Commission of IUB (NC-IUB), Newsletter. 1984. Archived from the original on 2012-10-15. Retrieved 2016-03-02.
↑ Suzuki N, Higuchi T, Nagano T (Aug 2002). "Multiple active intermediates in oxidation reaction catalyzed by synthetic heme-thiolate complex relevant to cytochrome p450". Journal of the American Chemical Society. 124 (32): 9622–8. doi:10.1021/ja0115013. PMID12167058.
↑ Cai J, Chen Y, Seth S, Furukawa S, Compans RW, Jones DP (Apr 2003). "Inhibition of influenza infection by glutathione". Free Radical Biology & Medicine. 34 (7): 928–36. doi:10.1016/S0891-5849(03)00023-6. PMID12654482.
↑ Ristoff E, Mayatepek E, Larsson A (Jul 2001). "Long-term clinical outcome in patients with glutathione synthetase deficiency". The Journal of Pediatrics. 139 (1): 79–84. doi:10.1067/mpd.2001.114480. PMID11445798.
↑ Jain A, Buist NR, Kennaway NG, Powell BR, Auld PA, Mårtensson J (Feb 1994). "Effect of ascorbate or N-acetylcysteine treatment in a patient with hereditary glutathione synthetase deficiency". The Journal of Pediatrics. 124 (2): 229–33. doi:10.1016/S0022-3476(94)70309-4. PMID8301428.
Law MY, Halliwell B (1986). "Purification and properties of glutathione synthetase from (Spinacia oleracea) leaves". Plant Sci. 43 (3): 185–191. doi:10.1016/0168-9452(86)90016-6.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.